

The Role of the Equine Herpesvirus Type 1 (EHV-1) US3-Encoded Protein Kinase in Actin Reorganization and Nuclear Egress
- PMID: 27754319
- PMCID: PMC5086611
- DOI: 10.3390/v8100275
The Role of the Equine Herpesvirus Type 1 (EHV-1) US3-Encoded Protein Kinase in Actin Reorganization and Nuclear Egress
Abstract
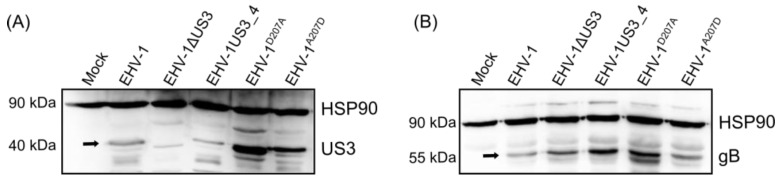
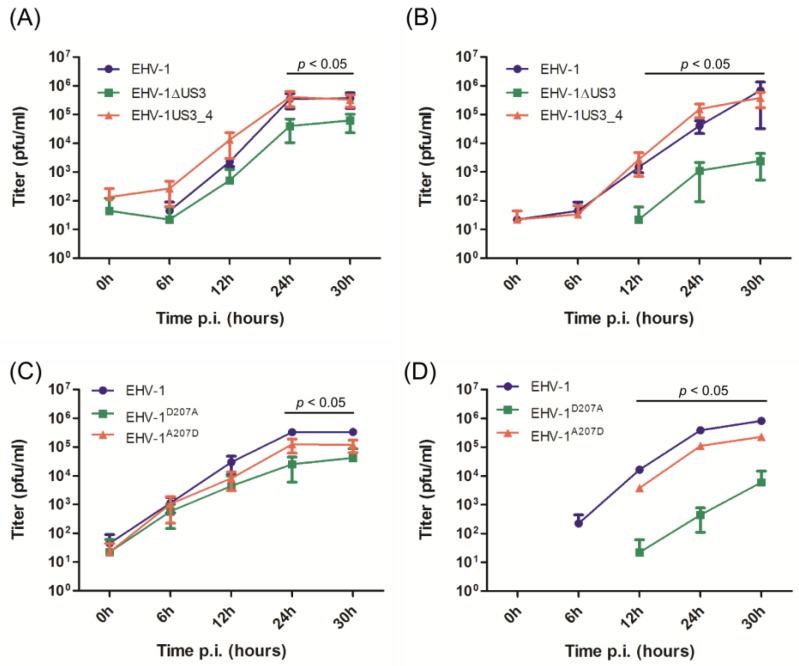
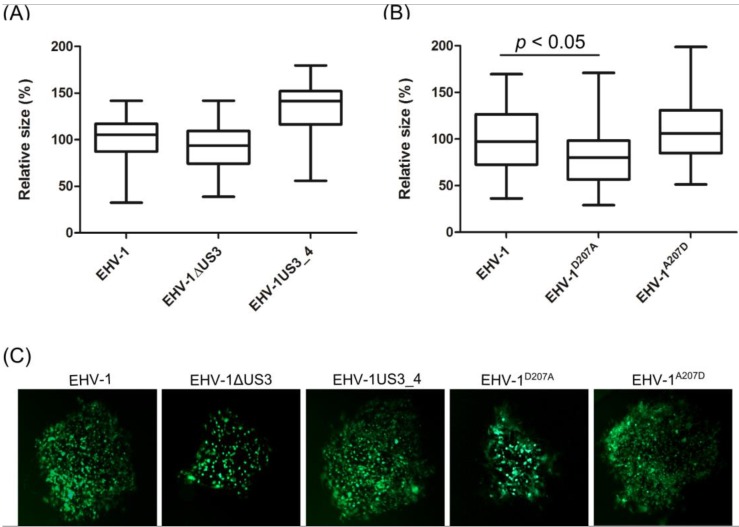
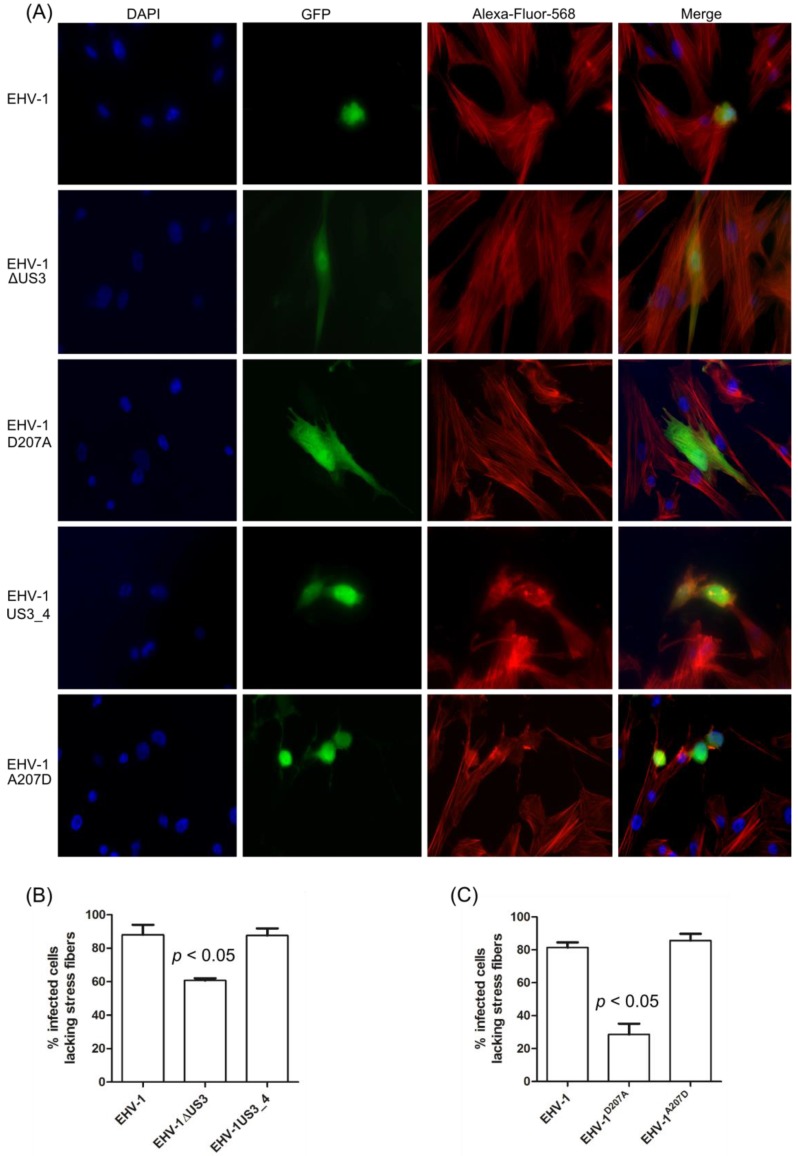
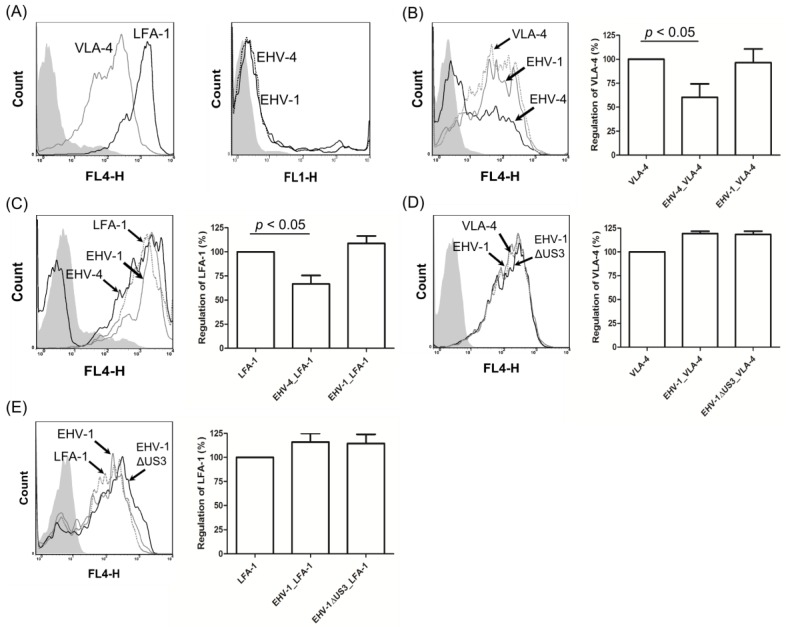
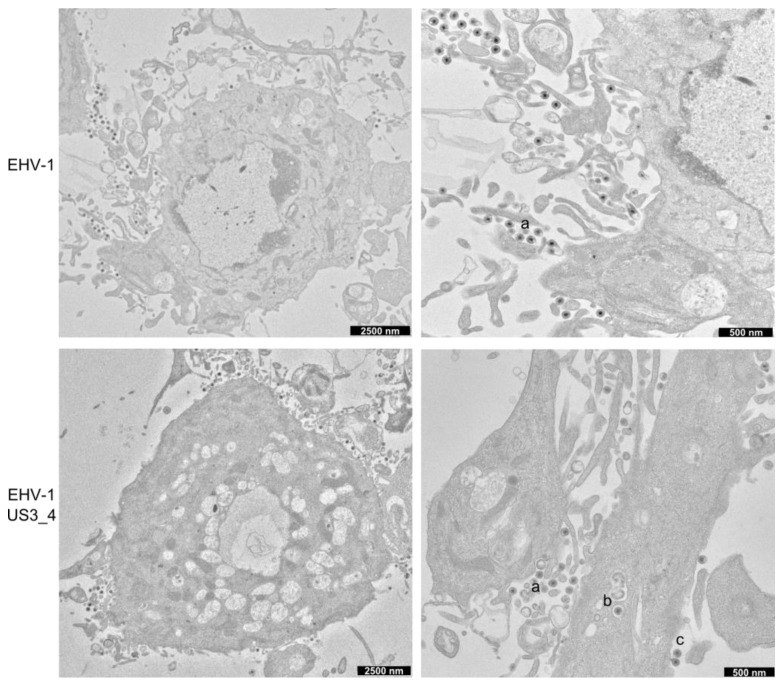
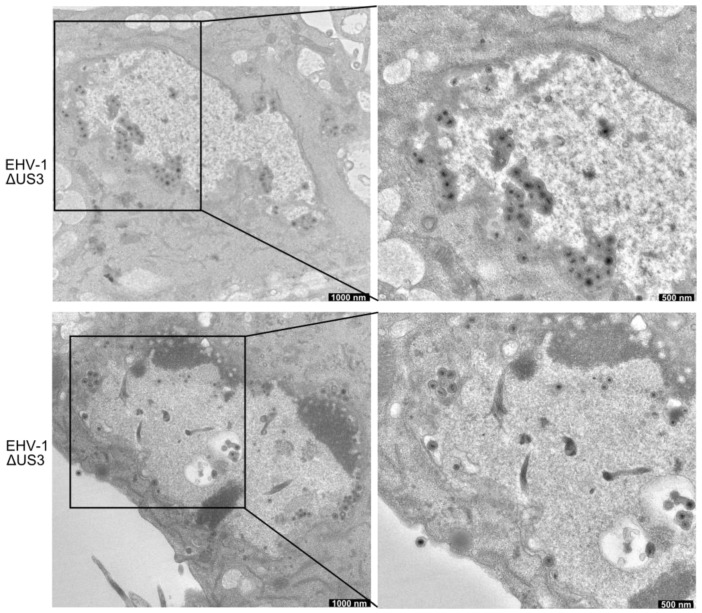
The serine-threonine protein kinase encoded by US3 gene (pUS3) of alphaherpesviruses was shown to modulate actin reorganization, cell-to-cell spread, and virus egress in a number of virus species. However, the role of the US3 orthologues of equine herpesvirus type 1 and 4 (EHV-1 and EHV-4) has not yet been studied. Here, we show that US3 is not essential for virus replication in vitro. However, growth rates and plaque diameters of a US3-deleted EHV-1 and a mutant in which the catalytic active site was destroyed were significantly reduced when compared with parental and revertant viruses or a virus in which EHV-1 US3 was replaced with the corresponding EHV-4 gene. The reduced plaque sizes were consistent with accumulation of primarily enveloped virions in the perinuclear space of the US3-negative EHV-1, a phenotype that was also rescued by the EHV-4 orthologue. Furthermore, actin stress fiber disassembly was significantly more pronounced in cells infected with parental EHV-1, revertant, or the recombinant EHV-1 expressing EHV-4 US3. Finally, we observed that deletion of US3 in EHV-1 did not affect the expression of adhesion molecules on the surface of infected cells.
Keywords: EHV-1; EHV-4; EM; US3; actin cytoskeleton; adhesion molecules.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Colle C.F., 3rd, Flowers C.C., O’Callaghan D.J. Open reading frames encoding a protein kinase, homolog of glycoprotein gX of pseudorabies virus, and a novel glycoprotein map within the unique short segment of equine herpesvirus type 1. Virology. 1992;188:545–557. doi: 10.1016/0042-6822(92)90509-N. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
