

C-C Chemokine Receptor Type 2-Dependent Migration of Myeloid-Derived Suppressor Cells in Protection of Islet Transplants
- PMID: 27755503
- PMCID: PMC5393972
- DOI: 10.1097/TP.0000000000001529
C-C Chemokine Receptor Type 2-Dependent Migration of Myeloid-Derived Suppressor Cells in Protection of Islet Transplants
Abstract
Background: Islet transplantation is a promising therapeutic approach to restore the physical response to blood glucose in type 1 diabetes. Current chronic use of immunosuppressive reagents for preventing islet allograft rejection is associated with severe complications. In addition, many of the immunosuppressive drugs are diabetogenic. The induction of transplant tolerance to eliminate the dependency on immunosuppression is ideal, but remains challenging.
Methods: Addition of hepatic stellate cells allowed generation of myeloid-derived suppressor cells (MDSC) from precursors in mouse bone marrow. Migration of MDSC was examined in an islet allograft transplant model by tracking the systemic administered MDSC from CD45.1 congenic mice.
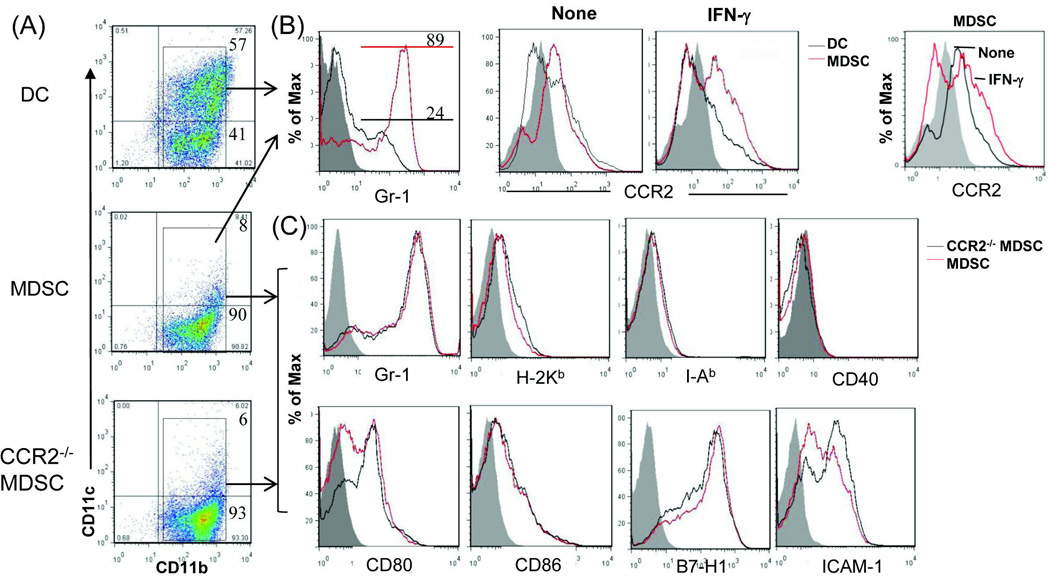
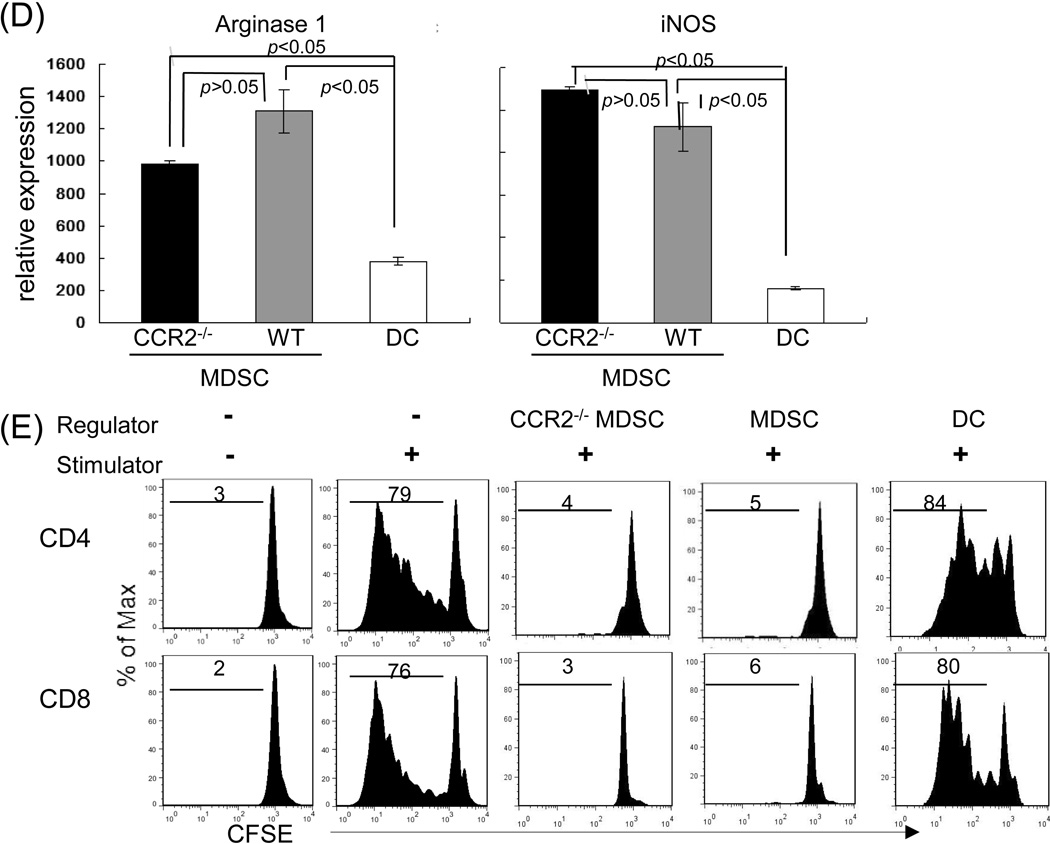
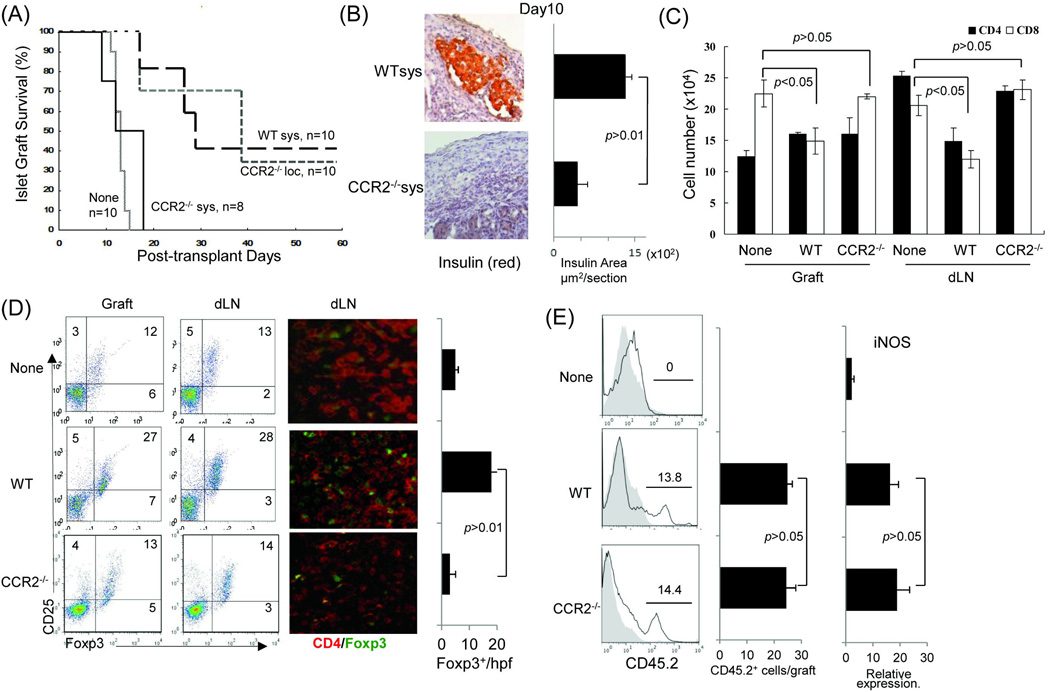
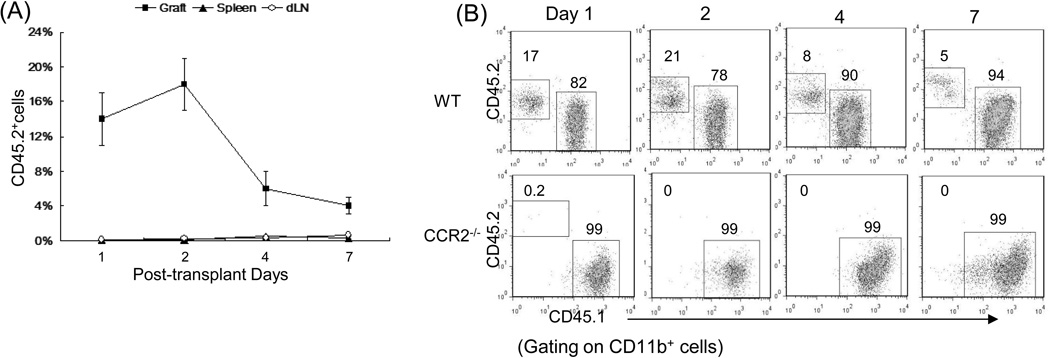
Results: The generated MDSC were expressed C-C chemokine receptor type 2 (CCR2), which was enhanced by exposure to interferon-γ. A single systemic administration of MDSC markedly prolonged survival of islet allografts without requirement of immunosuppression. Tracking the administered MDSC showed that they promptly migrated to the islet graft sites, at which point they exerted potent immune suppressive activity by inhibiting CD8 T cells, enhancing regulatory T cell activity. MDSC generated from CCR2 mice failed to be mobilized and lost tolerogenic activity in vivo, but sustained suppressive activity in vitro.
Conclusions: MDSC migration was dependent on expression of CCR2, whereas CCR2 does not directly participate in immune suppression. Expression of CCR2 needs to be closely monitored for quality control purpose when MDSC are generated in vitro for immune therapy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Tolerogenic Role of Myeloid Suppressor Cells in Organ Transplantation.Front Immunol. 2019 Mar 6;10:374. doi: 10.3389/fimmu.2019.00374. eCollection 2019. Front Immunol. 2019. PMID: 30894860 Free PMC article. Review.
-
Blocking the monocyte chemoattractant protein-1/CCR2 chemokine pathway induces permanent survival of islet allografts through a programmed death-1 ligand-1-dependent mechanism.J Immunol. 2003 Dec 15;171(12):6929-35. doi: 10.4049/jimmunol.171.12.6929. J Immunol. 2003. PMID: 14662900
-
The mTOR Deficiency in Monocytic Myeloid-Derived Suppressor Cells Protects Mouse Cardiac Allografts by Inducing Allograft Tolerance.Front Immunol. 2021 Apr 9;12:661338. doi: 10.3389/fimmu.2021.661338. eCollection 2021. Front Immunol. 2021. PMID: 33897705 Free PMC article.
-
Myeloid-derived suppressor cells protect islet transplants by B7-H1 mediated enhancement of T regulatory cells.Transplantation. 2012 Feb 15;93(3):272-82. doi: 10.1097/TP.0b013e31823ffd39. Transplantation. 2012. PMID: 22179405 Free PMC article.
-
Chemokines and their receptors in islet allograft rejection and as targets for tolerance induction.Cell Transplant. 2006;15(4):295-309. Cell Transplant. 2006. PMID: 16898223 Review.
Cited by
-
Myeloid-derived suppressor cells increase and inhibit donor-reactive T cell responses to graft intestinal epithelium in intestinal transplant patients.Am J Transplant. 2018 Oct;18(10):2544-2558. doi: 10.1111/ajt.14718. Epub 2018 Apr 17. Am J Transplant. 2018. PMID: 29509288 Free PMC article.
-
Recent advances in myeloid-derived suppressor cell biology.Front Med. 2021 Apr;15(2):232-251. doi: 10.1007/s11684-020-0797-2. Epub 2020 Sep 2. Front Med. 2021. PMID: 32876877 Review.
-
Tolerogenic Role of Myeloid Suppressor Cells in Organ Transplantation.Front Immunol. 2019 Mar 6;10:374. doi: 10.3389/fimmu.2019.00374. eCollection 2019. Front Immunol. 2019. PMID: 30894860 Free PMC article. Review.
-
Preliminary assessment of the feasibility of autologous myeloid-derived suppressor cell infusion in non-human primate kidney transplantation.Transpl Immunol. 2019 Oct;56:101225. doi: 10.1016/j.trim.2019.101225. Epub 2019 Jul 19. Transpl Immunol. 2019. PMID: 31330261 Free PMC article.
-
Adoptive Transfer of Regulatory Immune Cells in Organ Transplantation.Front Immunol. 2021 Mar 2;12:631365. doi: 10.3389/fimmu.2021.631365. eCollection 2021. Front Immunol. 2021. PMID: 33737934 Free PMC article. Review.
References
-
- Barr CC. Retinopathy and nephropathy in patients with type 1 diabetes four years after a trial of intensive insulin therapy, by The Diabetes Control and Complications Trial/Epidemiology of Diabetes Interventions and Complications Research Group. N. Engl. J. Med. 2000;342:381–389. - PubMed
-
- Senior P, Kin T, Shapior J, Koh A. Islet transplantation at the university of alberta: Status update and review of progress over the last decade. Can J Diabetes. 2012;36:32–37.
-
- Gala-Lopez B, Pepper AR, Shapiro AM. Biologic agents in islet transplantation. Curr Diab Rep. 2013;13:713–722. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
