

Using C. elegans Forward and Reverse Genetics to Identify New Compounds with Anthelmintic Activity
- PMID: 27755544
- PMCID: PMC5068747
- DOI: 10.1371/journal.pntd.0005058
Using C. elegans Forward and Reverse Genetics to Identify New Compounds with Anthelmintic Activity
Abstract
Background: The lack of new anthelmintic agents is of growing concern because it affects human health and our food supply, as both livestock and plants are affected. Two principal factors contribute to this problem. First, nematode resistance to anthelmintic drugs is increasing worldwide and second, many effective nematicides pose environmental hazards. In this paper we address this problem by deploying a high throughput screening platform for anthelmintic drug discovery using the nematode Caenorhabditis elegans as a surrogate for infectious nematodes. This method offers the possibility of identifying new anthelmintics in a cost-effective and timely manner.
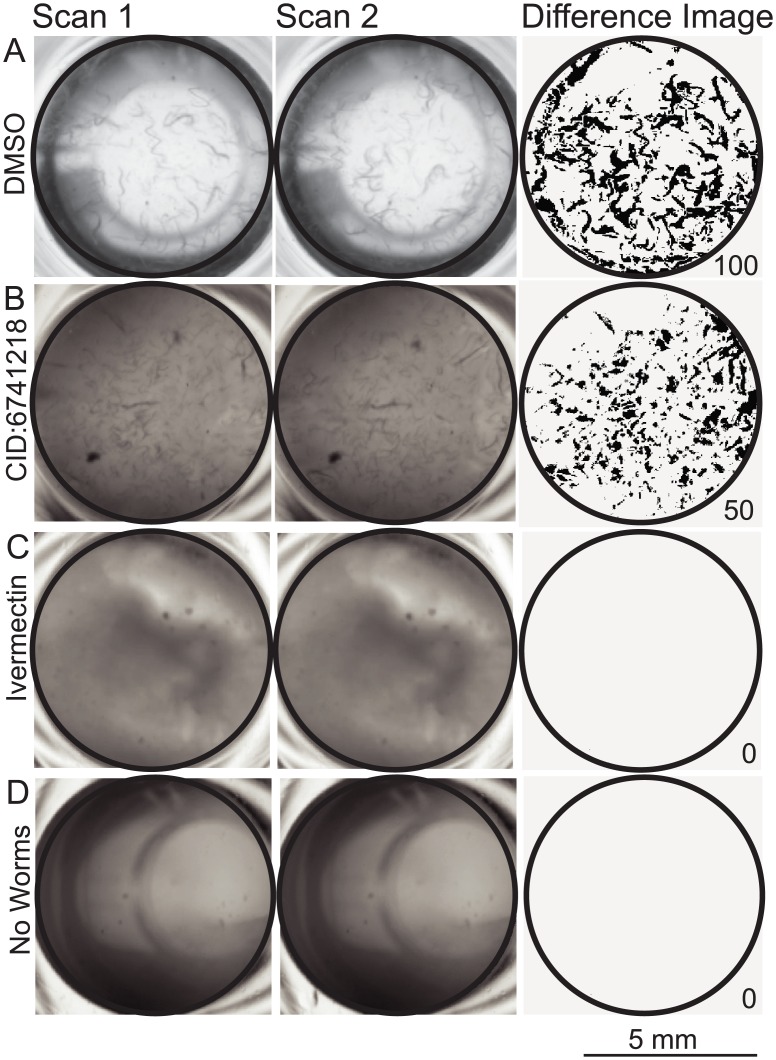
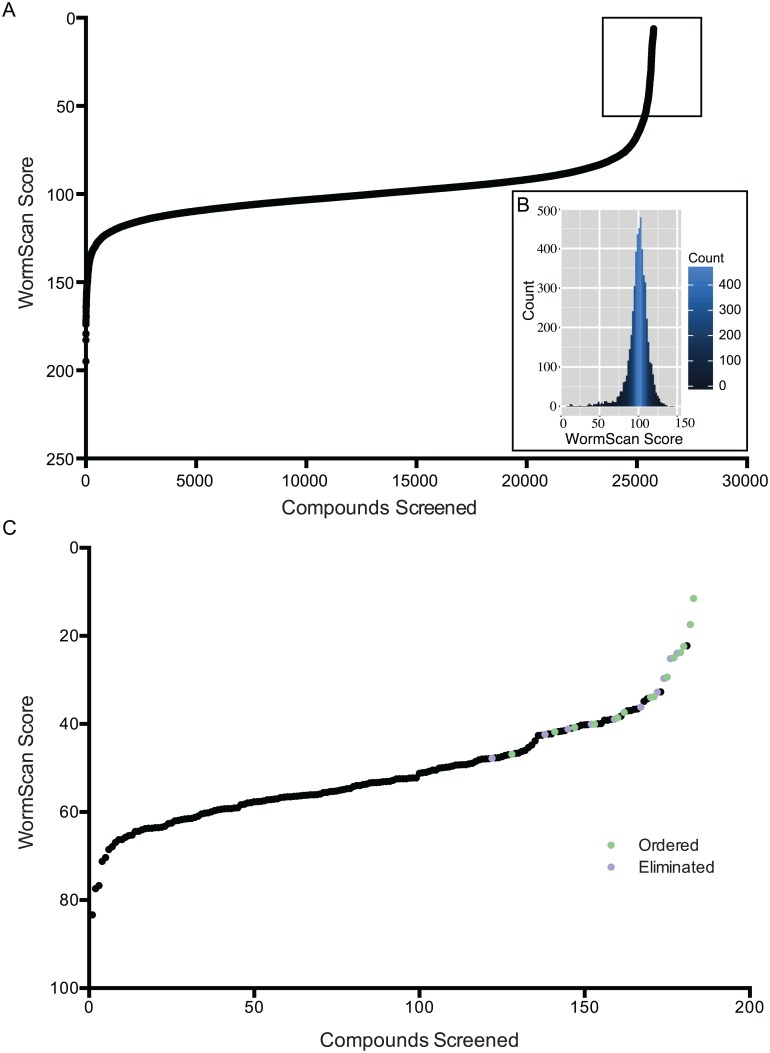
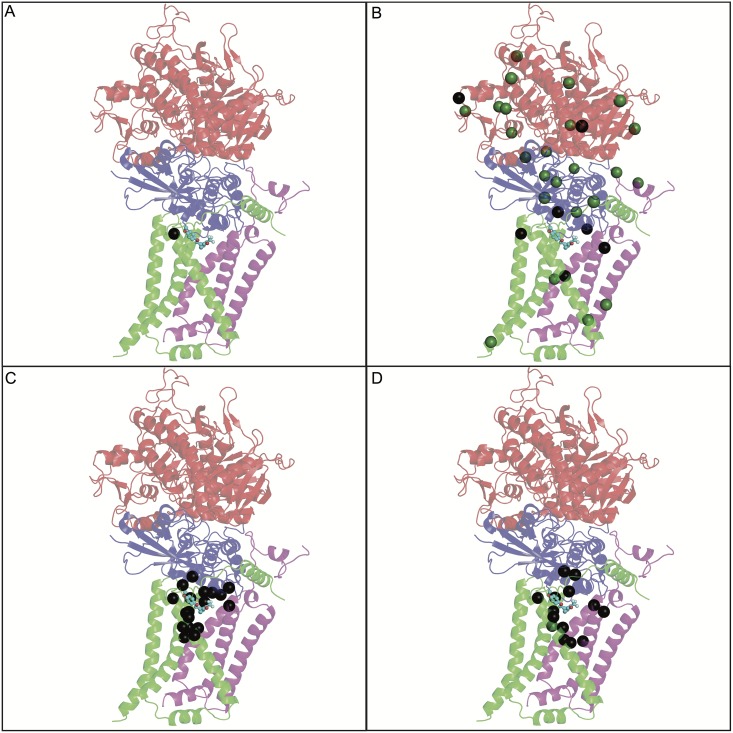
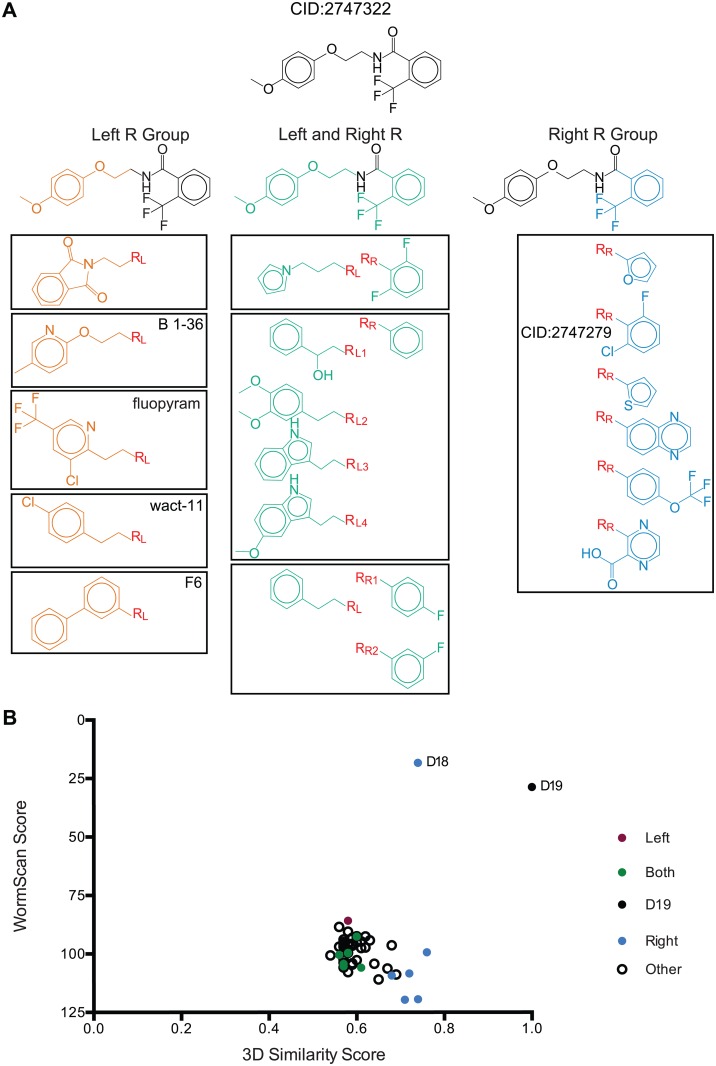
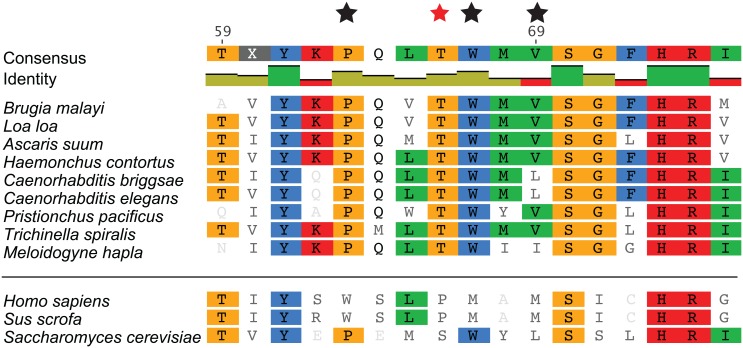
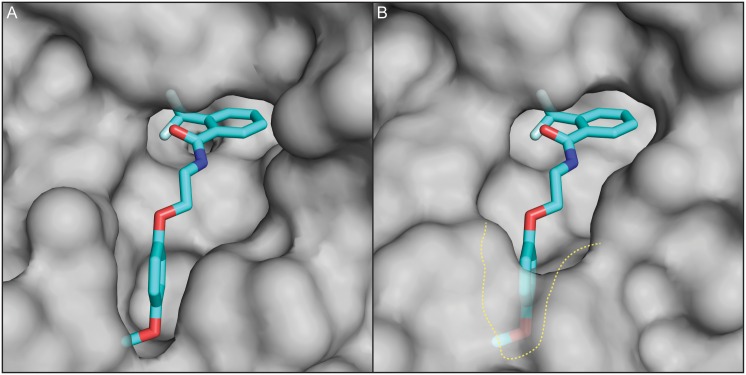
Methods/principal findings: Using our high throughput screening platform we have identified 14 new potential anthelmintics by screening more than 26,000 compounds from the Chembridge and Maybridge chemical libraries. Using phylogenetic profiling we identified a subset of the 14 compounds as potential anthelmintics based on the relative sensitivity of C. elegans when compared to yeast and mammalian cells in culture. We showed that a subset of these compounds might employ mechanisms distinct from currently used anthelmintics by testing diverse drug resistant strains of C. elegans. One of these newly identified compounds targets mitochondrial complex II, and we used structural analysis of the target to suggest how differential binding of this compound may account for its different effects in nematodes versus mammalian cells.
Conclusions/significance: The challenge of anthelmintic drug discovery is exacerbated by several factors; including, 1) the biochemical similarity between host and parasite genomes, 2) the geographic location of parasitic nematodes and 3) the rapid development of resistance. Accordingly, an approach that can screen large compound collections rapidly is required. C. elegans as a surrogate parasite offers the ability to screen compounds rapidly and, equally importantly, with specificity, thus reducing the potential toxicity of these compounds to the host and the environment. We believe this approach will help to replenish the pipeline of potential nematicides.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Anthelmintic drugs and nematicides: studies in Caenorhabditis elegans.WormBook. 2014 Dec 16:1-29. doi: 10.1895/wormbook.1.143.2. WormBook. 2014. PMID: 25517625 Free PMC article. Review.
-
Deciphering the molecular determinants of cholinergic anthelmintic sensitivity in nematodes: When novel functional validation approaches highlight major differences between the model Caenorhabditis elegans and parasitic species.PLoS Pathog. 2018 May 2;14(5):e1006996. doi: 10.1371/journal.ppat.1006996. eCollection 2018 May. PLoS Pathog. 2018. PMID: 29719008 Free PMC article.
-
An automated high-throughput system for phenotypic screening of chemical libraries on C. elegans and parasitic nematodes.Int J Parasitol Drugs Drug Resist. 2018 Apr;8(1):8-21. doi: 10.1016/j.ijpddr.2017.11.004. Epub 2017 Dec 2. Int J Parasitol Drugs Drug Resist. 2018. PMID: 29223747 Free PMC article.
-
Discovery of genomic intervals that underlie nematode responses to benzimidazoles.PLoS Negl Trop Dis. 2018 Mar 30;12(3):e0006368. doi: 10.1371/journal.pntd.0006368. eCollection 2018 Mar. PLoS Negl Trop Dis. 2018. PMID: 29601575 Free PMC article.
-
Integrating metabolomics into the diagnosis and investigation of anthelmintic resistance.Trends Parasitol. 2024 Dec;40(12):1097-1106. doi: 10.1016/j.pt.2024.10.020. Epub 2024 Nov 20. Trends Parasitol. 2024. PMID: 39572328 Review.
Cited by
-
Structure-Based Bioisosterism Design, Synthesis, Biological Evaluation and In Silico Studies of Benzamide Analogs as Potential Anthelmintics.Molecules. 2022 Apr 20;27(9):2659. doi: 10.3390/molecules27092659. Molecules. 2022. PMID: 35566007 Free PMC article.
-
Anthelmintic drug actions in resistant and susceptible C. elegans revealed by electrophysiological recordings in a multichannel microfluidic device.Int J Parasitol Drugs Drug Resist. 2018 Dec;8(3):607-628. doi: 10.1016/j.ijpddr.2018.10.003. Epub 2018 Oct 30. Int J Parasitol Drugs Drug Resist. 2018. PMID: 30503202 Free PMC article.
-
A cell-based infection assay identifies efflux pump modulators that reduce bacterial intracellular load.PLoS Pathog. 2018 Jun 7;14(6):e1007115. doi: 10.1371/journal.ppat.1007115. eCollection 2018 Jun. PLoS Pathog. 2018. PMID: 29879224 Free PMC article.
-
A survey of the kinome pharmacopeia reveals multiple scaffolds and targets for the development of novel anthelmintics.Sci Rep. 2021 Apr 28;11(1):9161. doi: 10.1038/s41598-021-88150-6. Sci Rep. 2021. PMID: 33911106 Free PMC article.
-
Biochemometry identifies ostruthin as pluripotent antimicrobial and anthelmintic agent from masterwort.iScience. 2023 Aug 3;26(9):107523. doi: 10.1016/j.isci.2023.107523. eCollection 2023 Sep 15. iScience. 2023. PMID: 37636068 Free PMC article.
References
-
- Organization WH. Deworming for health and development: report of the Third Global Meeting of the Partners for Parasite Control 2005.
-
- Hassan MA, Pham TH, Shi H. Nematodes threats to global food security. Acta Agriculturae Scandinavica, Section B—Soil & Plant Science. 2013. 5048972262744344578related:AkituYWNEUYJ. 10.1080/09064710.2013.794858 - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
