

Growth and maturation of heart valves leads to changes in endothelial cell distribution, impaired function, decreased metabolism and reduced cell proliferation
- PMID: 27756541
- PMCID: PMC5154817
- DOI: 10.1016/j.yjmcc.2016.10.006
Growth and maturation of heart valves leads to changes in endothelial cell distribution, impaired function, decreased metabolism and reduced cell proliferation
Abstract
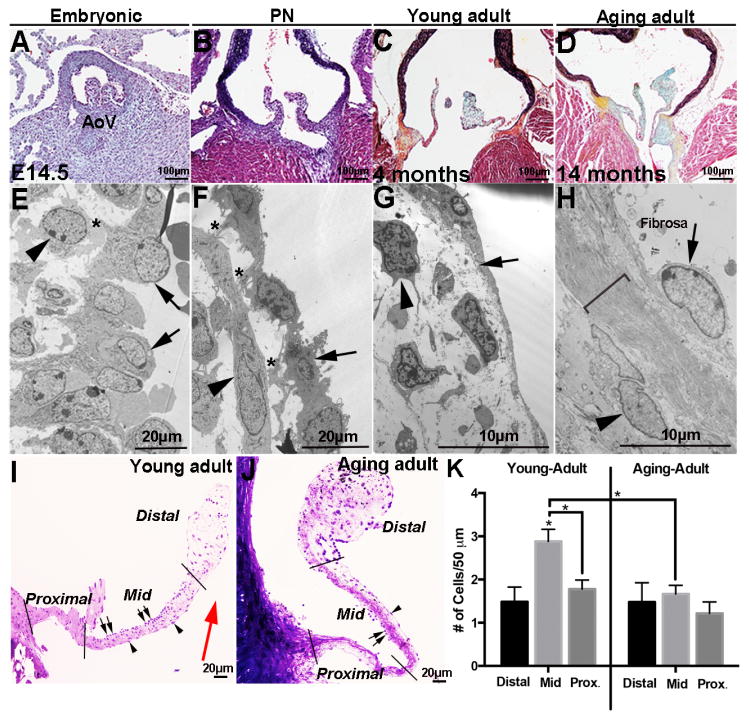
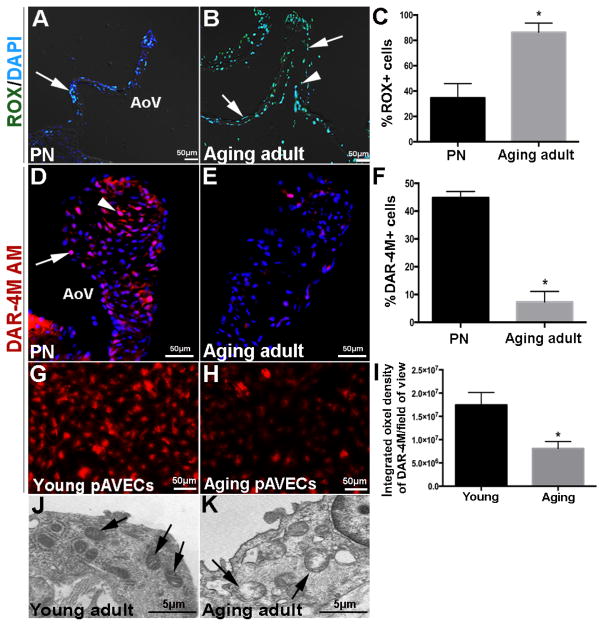
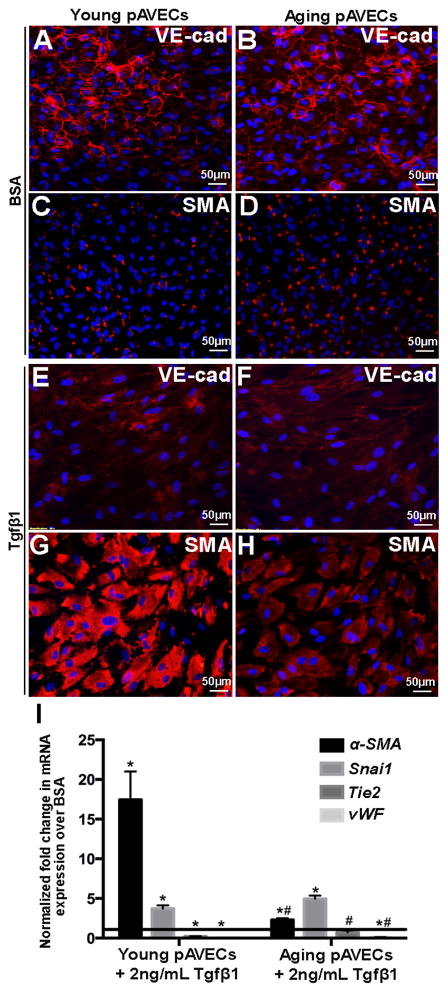
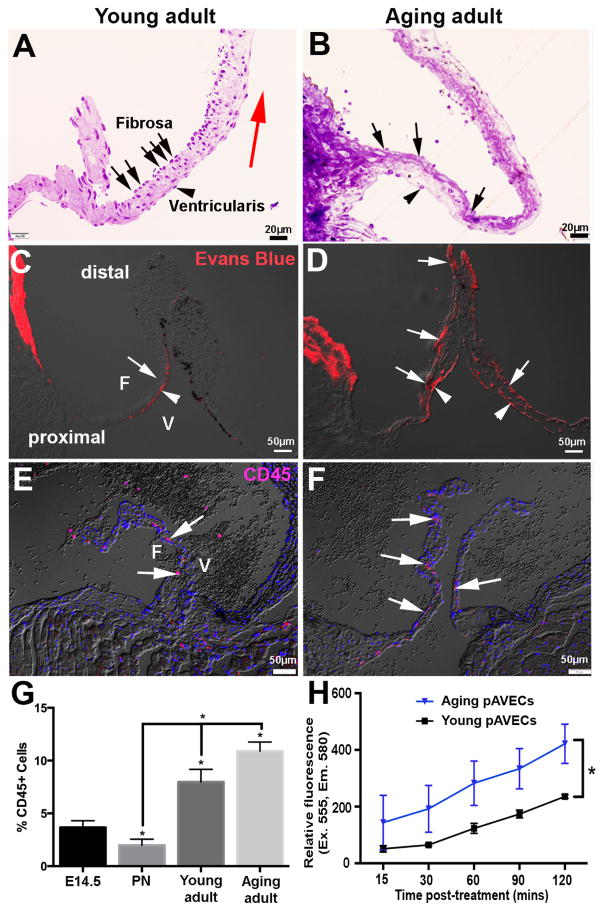
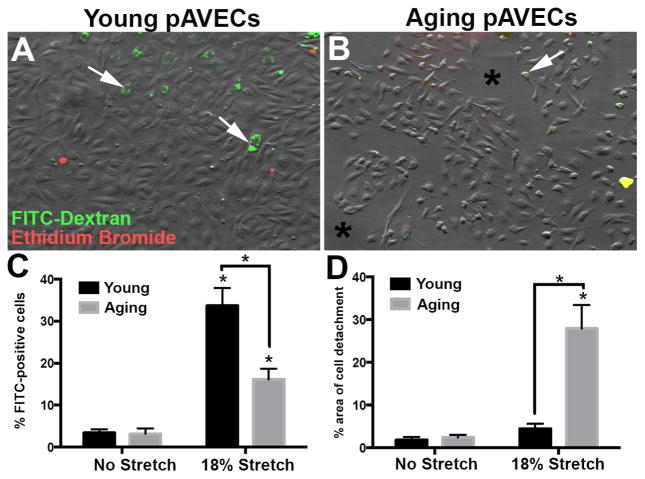
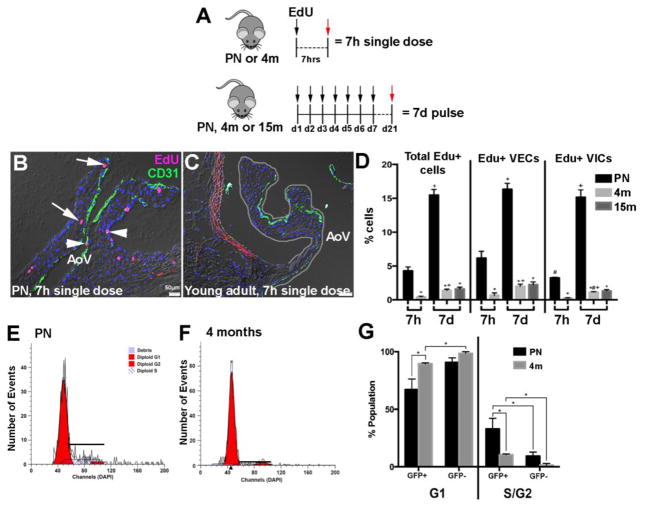
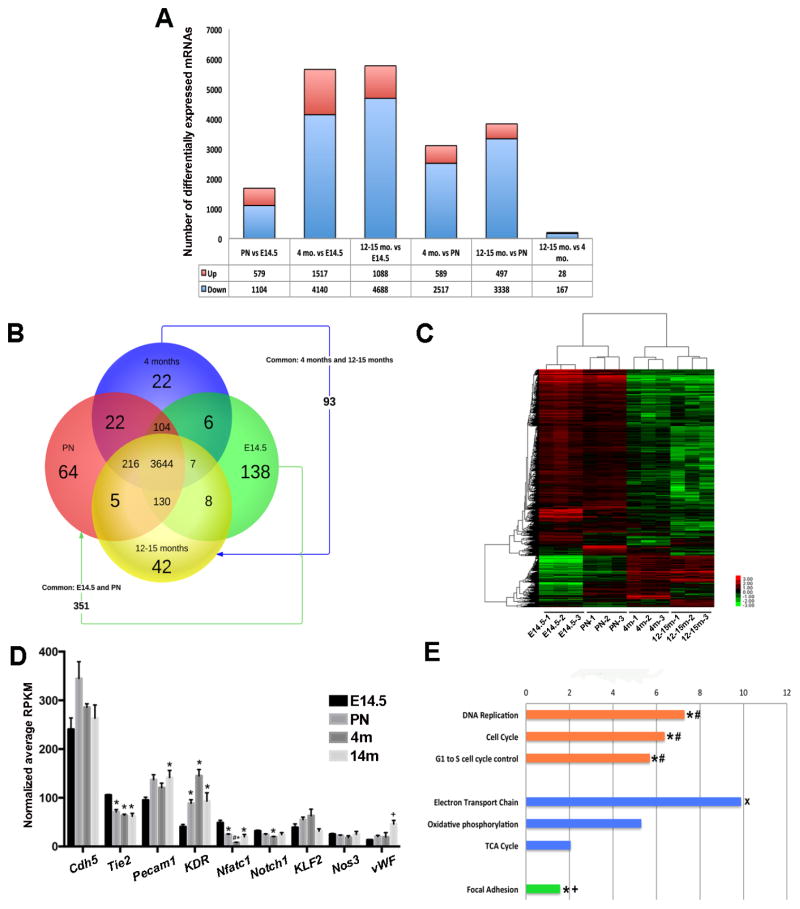
Risk factors of heart valve disease are well defined and prolonged exposure throughout life leads to degeneration and dysfunction in up to 33% of the population. While aortic valve replacement remains the most common need for cardiovascular surgery particularly in those aged over 65, the underlying mechanisms of progressive deterioration are unknown. In other cardiovascular systems, a decline in endothelial cell integrity and function play a major role in promoting pathological changes, and while similar mechanisms have been speculated in the valves, studies to support this are lacking. The goal of this study was to examine age-related changes in valve endothelial cell (VEC) distribution, morphology, function and transcriptomes during critical stages of valve development (embryonic), growth (postnatal (PN)), maintenance (young adult) and aging (aging adult). Using a combination of in vivo mouse, and in vitro porcine assays we show that VEC function including, nitric oxide bioavailability, metabolism, endothelial-to-mesenchymal potential, membrane self-repair and proliferation decline with age. In addition, density of VEC distribution along the endothelium decreases and this is associated with changes in morphology, decreased cell-cell interactions, and increased permeability. These changes are supported by RNA-seq analysis showing that focal adhesion-, cell cycle-, and oxidative phosphorylation-associated biological processes are negatively impacted by aging. Furthermore, by performing high-throughput analysis we are able to report the differential and common transcriptomes of VECs at each time point that can provide insights into the mechanisms underlying age-related dysfunction. These studies suggest that maturation of heart valves over time is a multifactorial process and this study has identified several key parameters that may contribute to impairment of the valve to maintain critical structure-function relationships; leading to degeneration and disease.
Keywords: Endothelial cell; Growth; Heart valve; Maturation.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Hinton RB, Jr, Lincoln J, Deutsch GH, Osinska H, Manning PB, Benson DW, et al. Extracellular matrix remodeling and organization in developing and diseased aortic valves. Circ Res. 2006;98(11):1431–8. - PubMed
-
- Tao G, Kotick JD, Lincoln J. Heart valve development, maintenance, and disease: the role of endothelial cells. Curr Top Dev Biol. 2012;100:203–32. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
