

Oxidized Guanine Base Lesions Function in 8-Oxoguanine DNA Glycosylase-1-mediated Epigenetic Regulation of Nuclear Factor κB-driven Gene Expression
- PMID: 27756845
- PMCID: PMC5207254
- DOI: 10.1074/jbc.M116.751453
Oxidized Guanine Base Lesions Function in 8-Oxoguanine DNA Glycosylase-1-mediated Epigenetic Regulation of Nuclear Factor κB-driven Gene Expression
Abstract
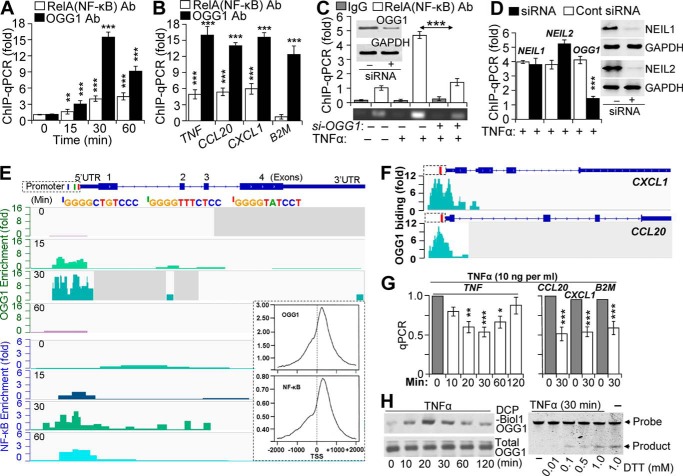
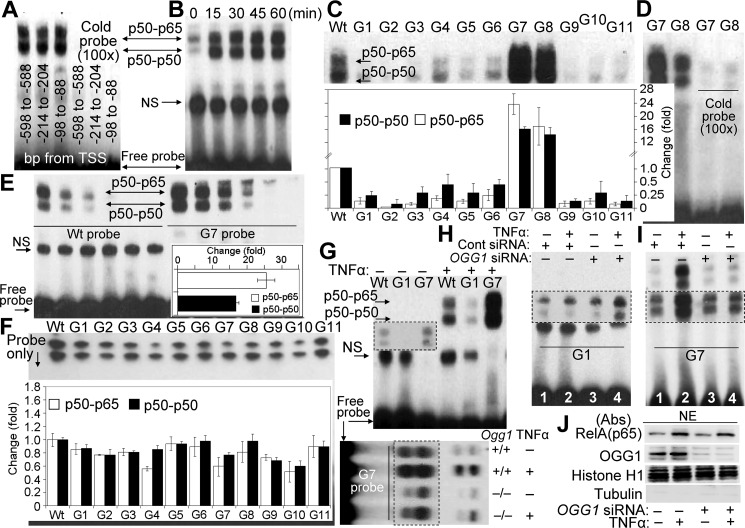
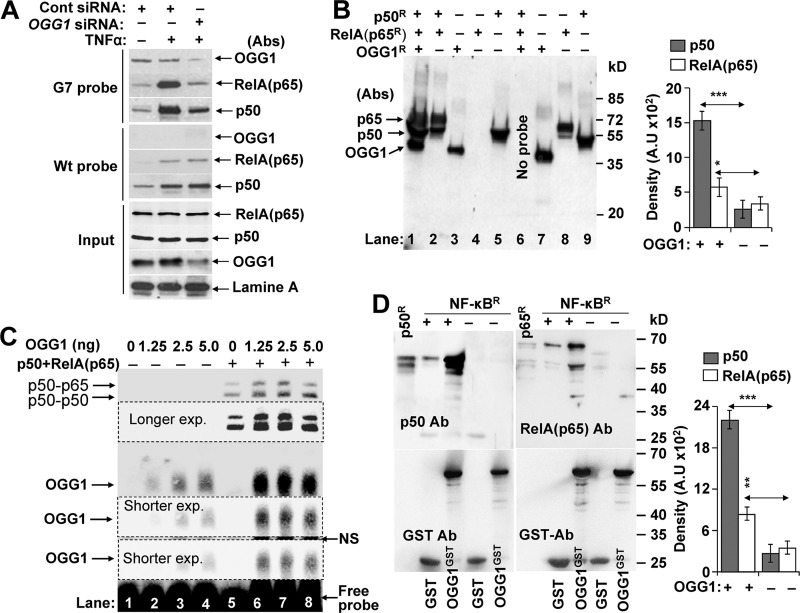
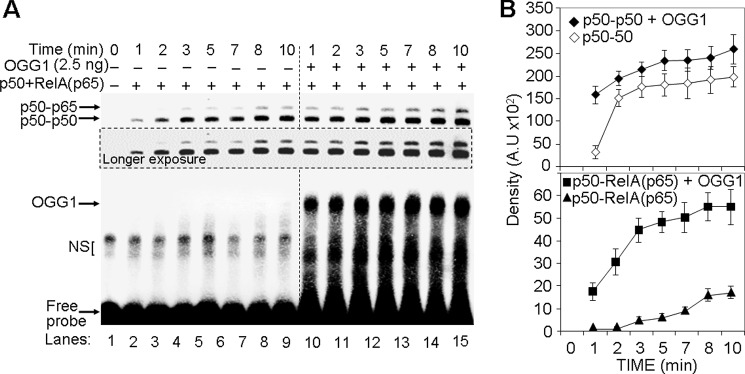
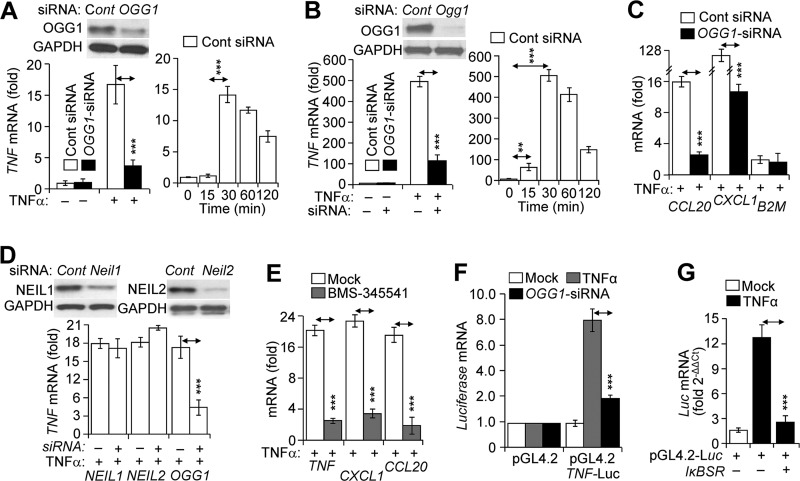
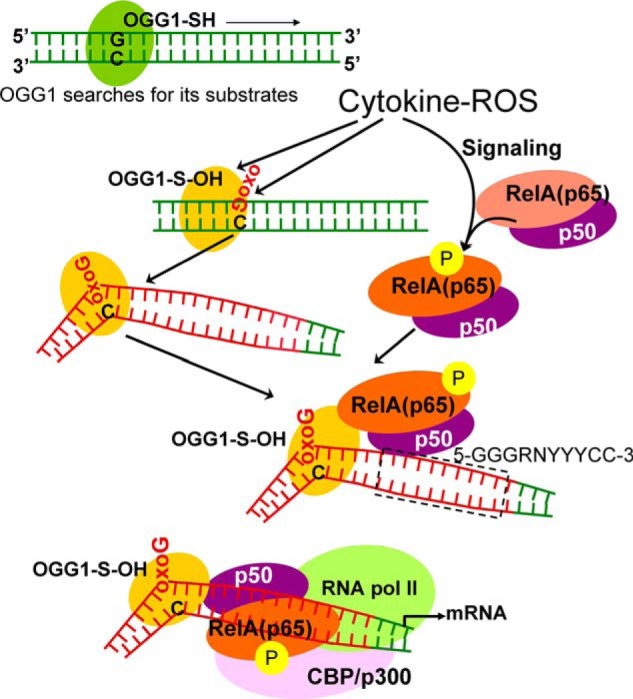
A large percentage of redox-responsive gene promoters contain evolutionarily conserved guanine-rich clusters; guanines are the bases most susceptible to oxidative modification(s). Consequently, 7,8-dihydro-8-oxoguanine (8-oxoG) is one of the most abundant base lesions in promoters and is primarily repaired via the 8-oxoguanine DNA glycosylase-1 (OOG1)-initiated base excision repair pathway. In view of a prompt cellular response to oxidative challenge, we hypothesized that the 8-oxoG lesion and the cognate repair protein OGG1 are utilized in transcriptional gene activation. Here, we document TNFα-induced enrichment of both 8-oxoG and OGG1 in promoters of pro-inflammatory genes, which precedes interaction of NF-κB with its DNA-binding motif. OGG1 bound to 8-oxoG upstream from the NF-κB motif increased its DNA occupancy by promoting an on-rate of both homodimeric and heterodimeric forms of NF-κB. OGG1 depletion decreased both NF-κB binding and gene expression, whereas Nei-like glycosylase-1 and -2 had a marginal effect. These results are the first to document a novel paradigm wherein the DNA repair protein OGG1 bound to its substrate is coupled to DNA occupancy of NF-κB and functions in epigenetic regulation of gene expression.
Keywords: 8-oxoguanine (8-oxoG); 8-oxoguanine glycosylase (OGG1); NF-κB transcription factor; promoter; reactive oxygen species (ROS).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Dizdaroglu M. (1992) Oxidative damage to DNA in mammalian chromatin. Mutat. Res. 275, 331–342 - PubMed
-
- Steenken S., and Jovanovic S. V. (1997) How easily oxidizable is DNA? One-electron reduction potentials of adenosine and guanosine radicals in aqueous solution. J. Am. Chem. Soc. 119, 617–618
-
- Mitra S., Hazra T. K., Roy R., Ikeda S., Biswas T., Lock J., Boldogh I., and Izumi T. (1997) Complexities of DNA base excision repair in mammalian cells. Mol. Cells 7, 305–312 - PubMed
-
- Lindahl T., and Barnes D. E. (2000) Repair of endogenous DNA damage. Cold Spring Harb. Symp. Quant. Biol. 65, 127–133 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
