

TIE2-mediated tyrosine phosphorylation of H4 regulates DNA damage response by recruiting ABL1
- PMID: 27757426
- PMCID: PMC5065225
- DOI: 10.1126/sciadv.1501290
TIE2-mediated tyrosine phosphorylation of H4 regulates DNA damage response by recruiting ABL1
Abstract
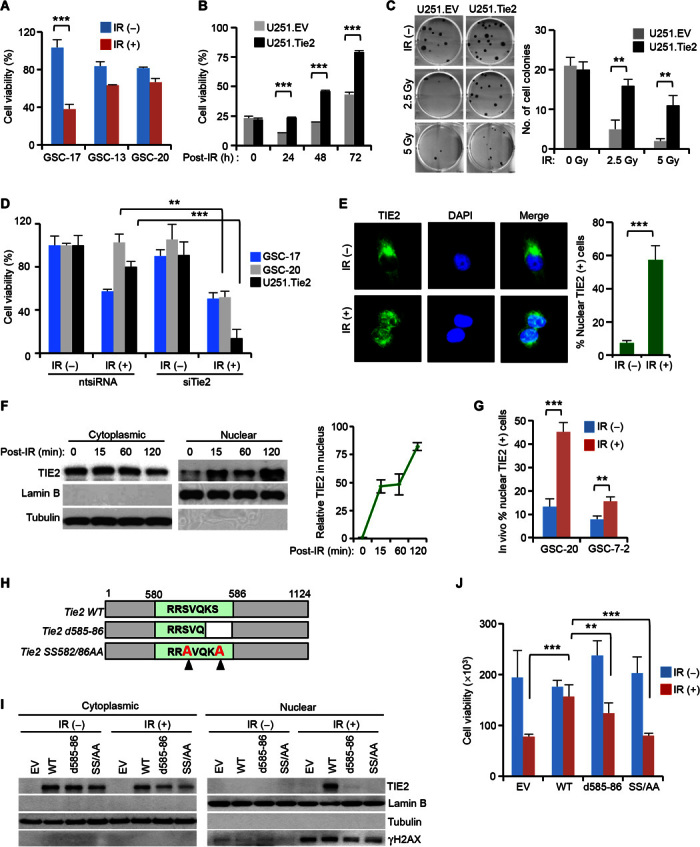
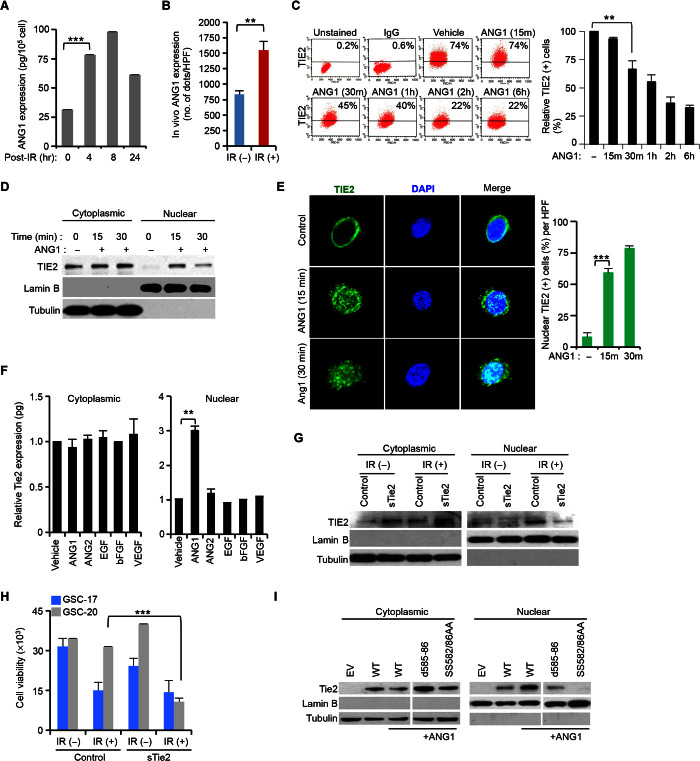
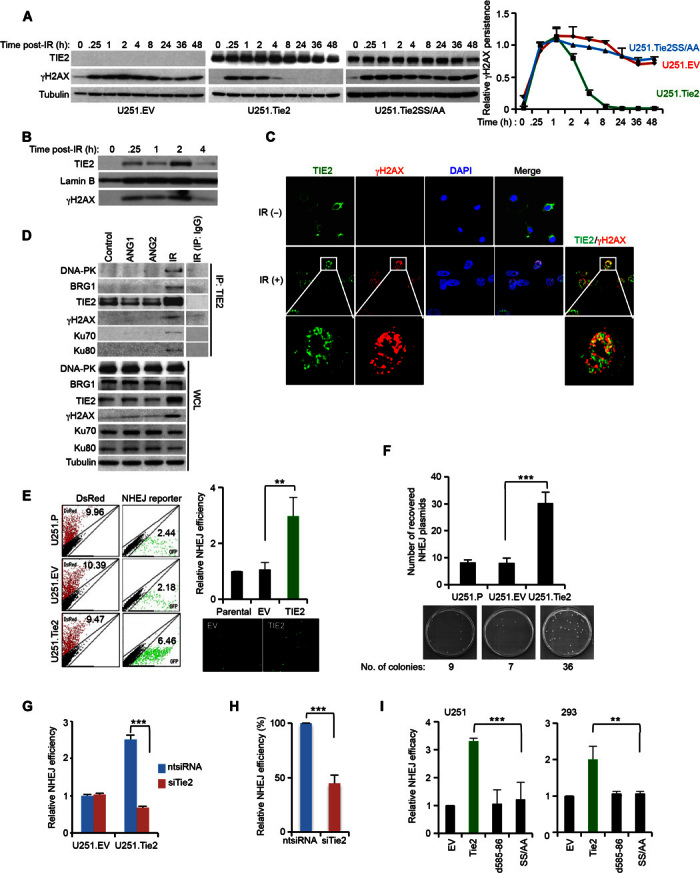
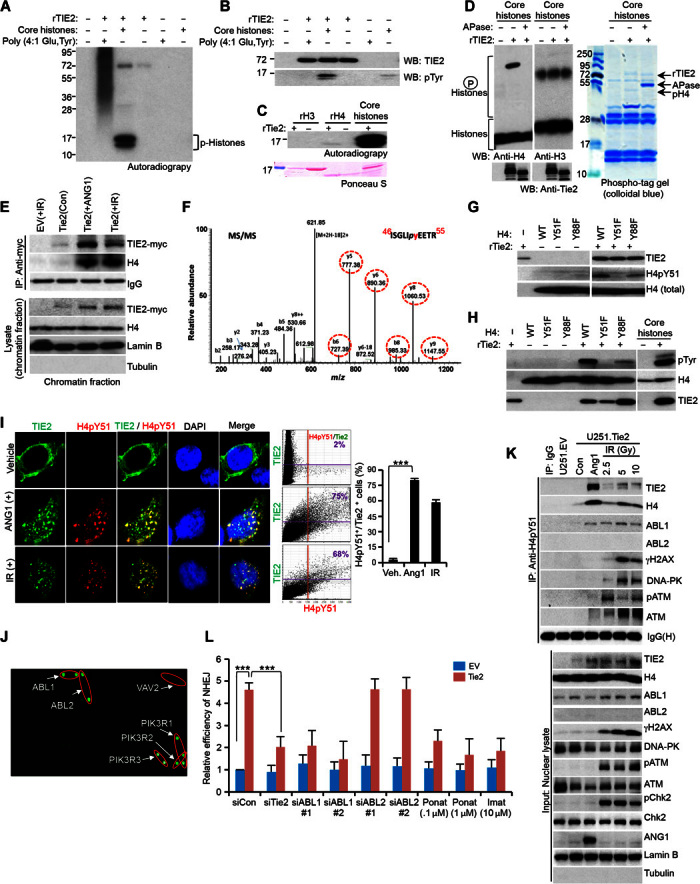
DNA repair pathways enable cancer cells to survive DNA damage induced after genotoxic therapies. Tyrosine kinase receptors (TKRs) have been reported as regulators of the DNA repair machinery. TIE2 is a TKR overexpressed in human gliomas at levels that correlate with the degree of increasing malignancy. Following ionizing radiation, TIE2 translocates to the nucleus, conferring cells with an enhanced nonhomologous end-joining mechanism of DNA repair that results in a radioresistant phenotype. Nuclear TIE2 binds to key components of DNA repair and phosphorylates H4 at tyrosine 51, which, in turn, is recognized by the proto-oncogene ABL1, indicating a role for nuclear TIE2 as a sensor for genotoxic stress by action as a histone modifier. H4Y51 constitutes the first tyrosine phosphorylation of core histones recognized by ABL1, defining this histone modification as a direct signal to couple genotoxic stress with the DNA repair machinery.
Keywords: ABL1; ANG1; Cell biology; DNA repair; NHEJ; TIE2; oncogenes.
Figures
Similar articles
-
TIE2 Associates with Caveolae and Regulates Caveolin-1 To Promote Their Nuclear Translocation.Mol Cell Biol. 2017 Oct 13;37(21):e00142-17. doi: 10.1128/MCB.00142-17. Print 2017 Nov 1. Mol Cell Biol. 2017. PMID: 28760776 Free PMC article.
-
c-Abl tyrosine kinase selectively regulates p73 nuclear matrix association.J Biol Chem. 2003 Sep 5;278(36):34475-82. doi: 10.1074/jbc.M301051200. Epub 2003 Jun 24. J Biol Chem. 2003. PMID: 12824179
-
Nuclear c-Abl-mediated tyrosine phosphorylation induces chromatin structural changes through histone modifications that include H4K16 hypoacetylation.Exp Cell Res. 2011 Dec 10;317(20):2874-903. doi: 10.1016/j.yexcr.2011.09.013. Epub 2011 Oct 2. Exp Cell Res. 2011. PMID: 22001646
-
Determination of cell fate by c-Abl activation in the response to DNA damage.Oncogene. 1998 Dec 24;17(25):3309-18. doi: 10.1038/sj.onc.1202571. Oncogene. 1998. PMID: 9916993 Review.
-
c‑Abl‑mediated tyrosine phosphorylation of DNA damage response proteins and implications in important cellular functions (Review).Mol Med Rep. 2020 Aug;22(2):612-619. doi: 10.3892/mmr.2020.11156. Epub 2020 May 18. Mol Med Rep. 2020. PMID: 32468042 Review.
Cited by
-
The role of caveolin-1 in tumors of the brain - functional and clinical implications.Cell Oncol (Dordr). 2019 Aug;42(4):423-447. doi: 10.1007/s13402-019-00447-x. Epub 2019 Apr 16. Cell Oncol (Dordr). 2019. PMID: 30993541 Review.
-
A high-throughput Gaussia luciferase reporter assay for screening potential gasdermin E activators against pancreatic cancer.Acta Pharm Sin B. 2023 Oct;13(10):4253-4272. doi: 10.1016/j.apsb.2023.07.018. Epub 2023 Jul 20. Acta Pharm Sin B. 2023. PMID: 37799380 Free PMC article.
-
Preserving genome integrity and function: the DNA damage response and histone modifications.Crit Rev Biochem Mol Biol. 2019 Jun;54(3):208-241. doi: 10.1080/10409238.2019.1620676. Epub 2019 Jun 4. Crit Rev Biochem Mol Biol. 2019. PMID: 31164001 Free PMC article. Review.
-
Normalizing Tumoral Vessels to Treat Cancer: An Out-of-the-Box Strategy Involving TIE2 Pathway.Transl Cancer Res. 2017 Mar;6(Suppl 2):S317-S320. doi: 10.21037/tcr.2017.03.64. Transl Cancer Res. 2017. PMID: 28944168 Free PMC article. No abstract available.
-
In silico molecular target prediction unveils mebendazole as a potent MAPK14 inhibitor.Mol Oncol. 2020 Dec;14(12):3083-3099. doi: 10.1002/1878-0261.12810. Epub 2020 Oct 18. Mol Oncol. 2020. PMID: 33021050 Free PMC article.
References
-
- Bao S., Wu Q., McLendon R. E., Hao Y., Shi Q., Hjelmeland A. B., Dewhirst M. W., Bigner D. D., Rich J. N., Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature 444, 756–760 (2006). - PubMed
-
- Hambardzumyan D., Squatrito M., Holland E. C., Radiation resistance and stem-like cells in brain tumors. Cancer Cell 10, 454–456 (2006). - PubMed
-
- Helleday T., Petermann E., Lundin C., Hodgson B., Sharma R. A., DNA repair pathways as targets for cancer therapy. Nat. Rev. Cancer 8, 193–204 (2008). - PubMed
-
- Lee O.-H., Xu J., Fueyo J., Fuller G. N., Aldape K. D., Alonso M. M., Piao Y., Liu T.-J., Lang F. F., Bekele B. N., Gomez-Manzano C., Expression of the receptor tyrosine kinase Tie2 in neoplastic glial cells is associated with integrin β1-dependent adhesion to the extracellular matrix. Mol. Cancer Res. 4, 915–926 (2006). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
