

Genome-Wide Mapping of Uncapped and Cleaved Transcripts Reveals a Role for the Nuclear mRNA Cap-Binding Complex in Cotranslational RNA Decay in Arabidopsis
- PMID: 27758893
- PMCID: PMC5134982
- DOI: 10.1105/tpc.16.00456
Genome-Wide Mapping of Uncapped and Cleaved Transcripts Reveals a Role for the Nuclear mRNA Cap-Binding Complex in Cotranslational RNA Decay in Arabidopsis
Abstract
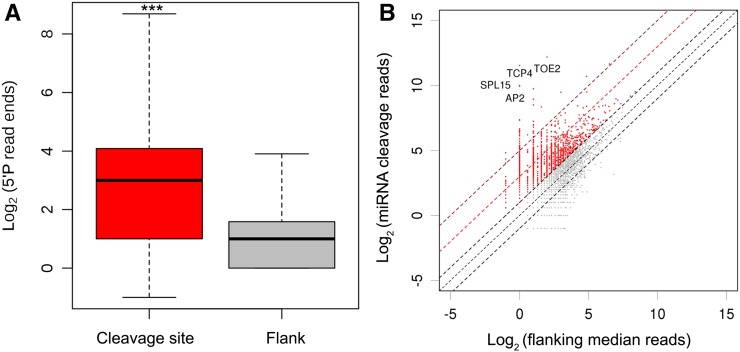
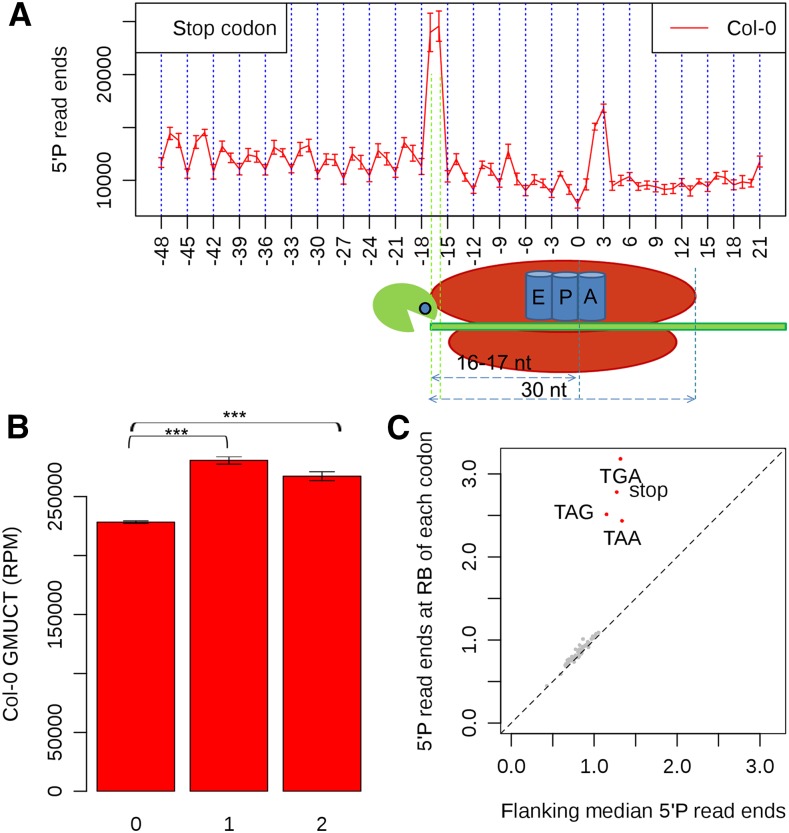
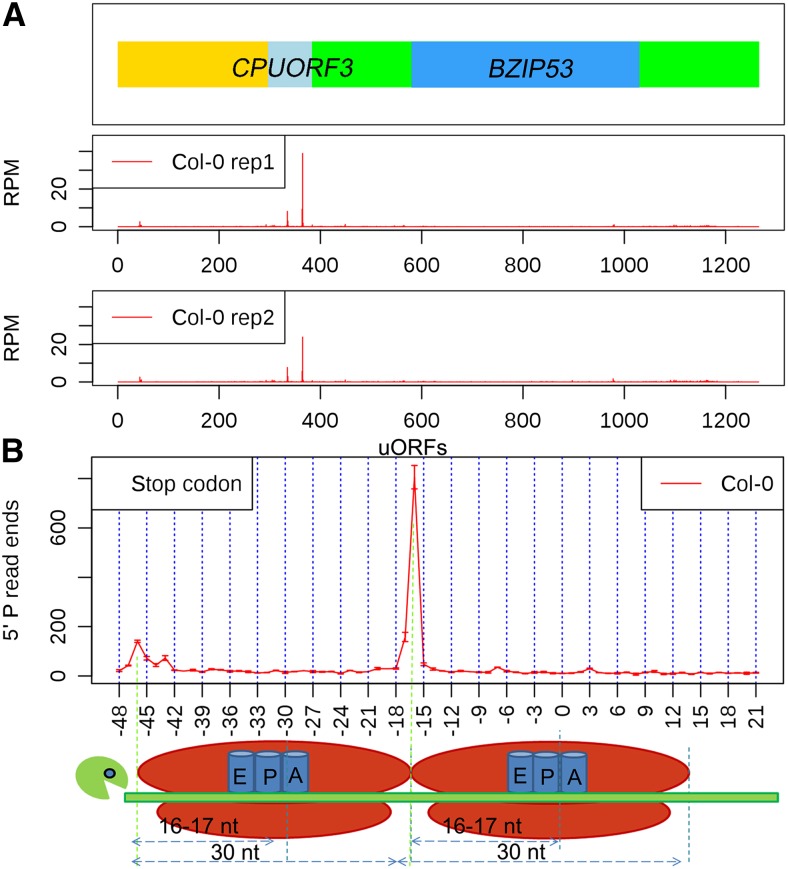
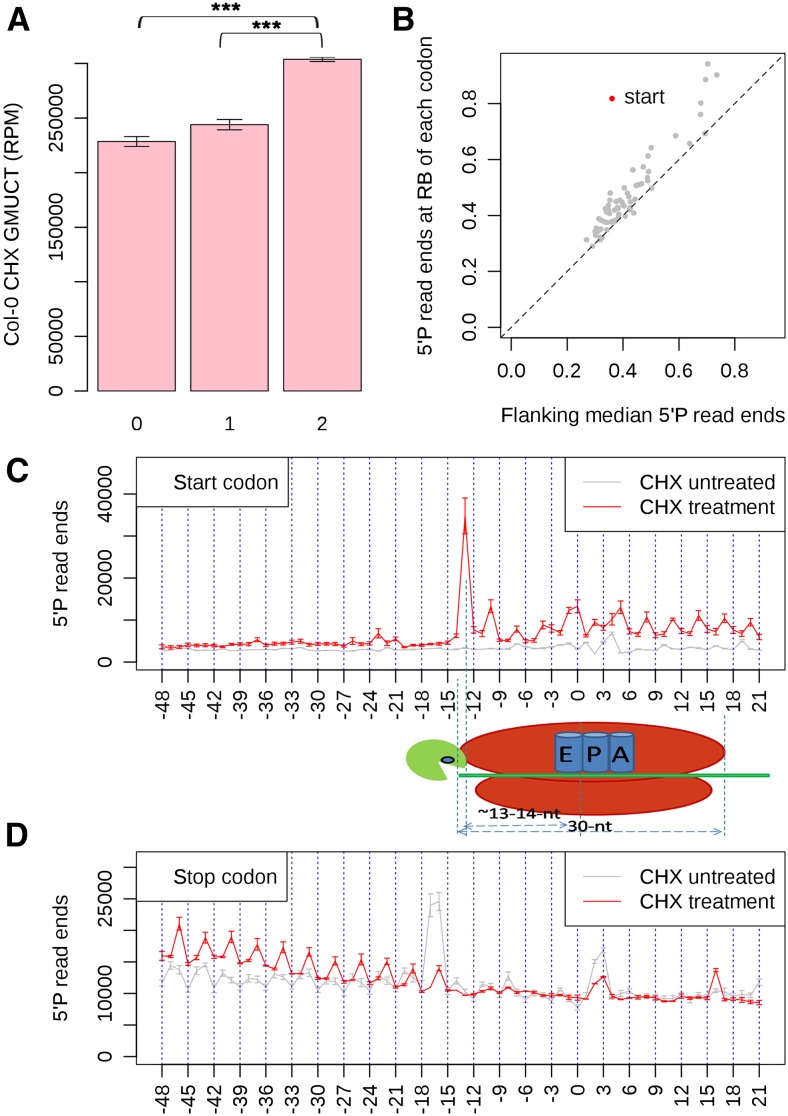
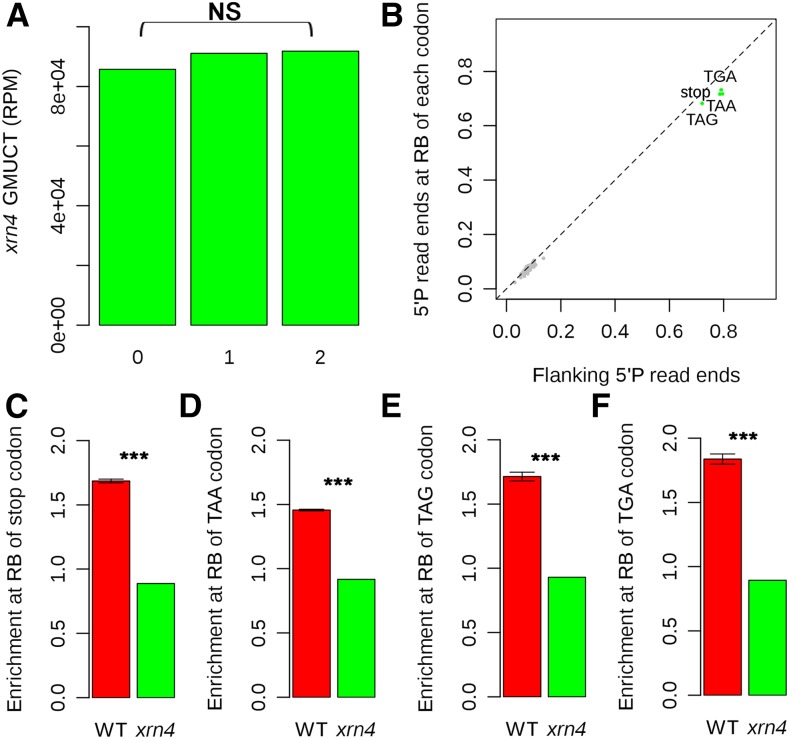
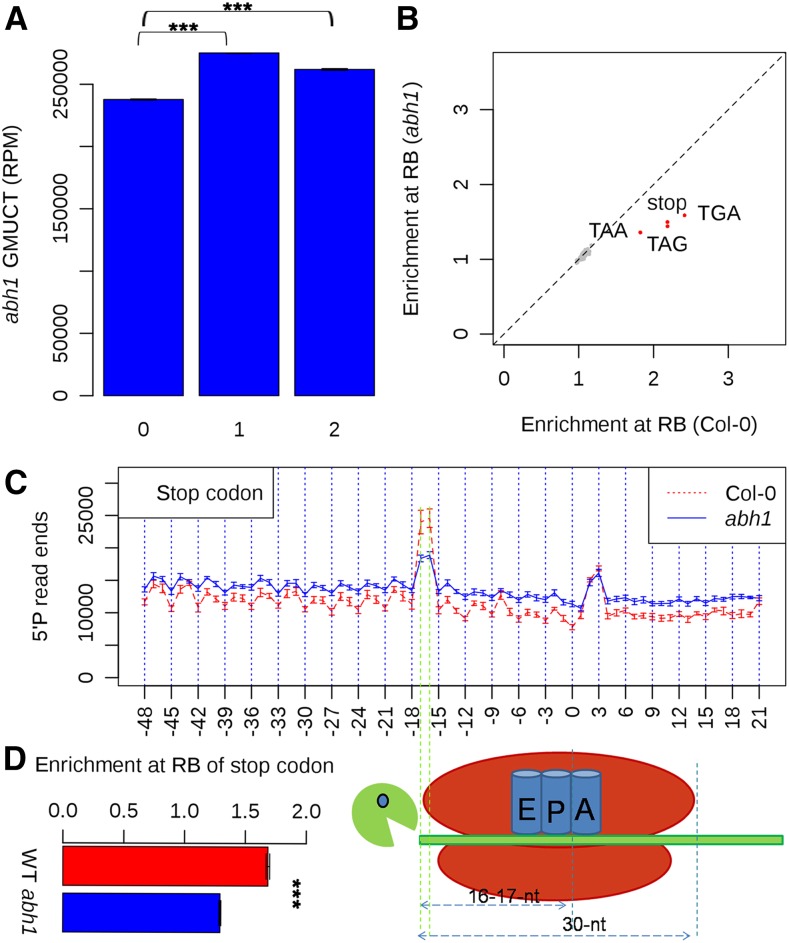
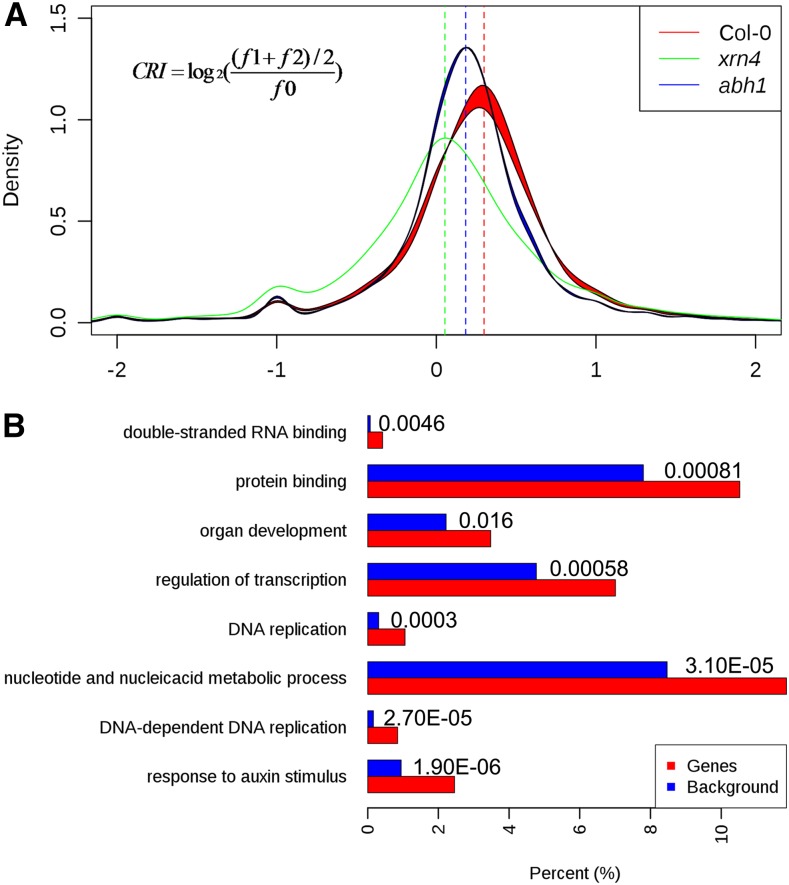
RNA turnover is necessary for controlling proper mRNA levels posttranscriptionally. In general, RNA degradation is via exoribonucleases that degrade RNA either from the 5' end to the 3' end, such as XRN4, or in the opposite direction by the multisubunit exosome complex. Here, we use genome-wide mapping of uncapped and cleaved transcripts to reveal the global landscape of cotranslational mRNA decay in the Arabidopsis thaliana transcriptome. We found that this process leaves a clear three nucleotide periodicity in open reading frames. This pattern of cotranslational degradation is especially evident near the ends of open reading frames, where we observe accumulation of cleavage events focused 16 to 17 nucleotides upstream of the stop codon because of ribosomal pausing during translation termination. Following treatment of Arabidopsis plants with the translation inhibitor cycloheximide, cleavage events accumulate 13 to 14 nucleotides upstream of the start codon where initiating ribosomes have been stalled with these sequences in their P site. Further analysis in xrn4 mutant plants indicates that cotranslational RNA decay is XRN4 dependent. Additionally, studies in plants lacking CAP BINDING PROTEIN80/ABA HYPERSENSITIVE1, the largest subunit of the nuclear mRNA cap binding complex, reveal a role for this protein in cotranslational decay. In total, our results demonstrate the global prevalence and features of cotranslational RNA decay in a plant transcriptome.
© 2016 American Society of Plant Biologists. All rights reserved.
Figures
Comment in
-
RNA Degradome Studies Give Insights into Ribosome Dynamics.Plant Cell. 2016 Oct;28(10):2348-2349. doi: 10.1105/tpc.16.00785. Epub 2016 Oct 17. Plant Cell. 2016. PMID: 27754877 Free PMC article. No abstract available.
Similar articles
-
Evidence that XRN4, an Arabidopsis homolog of exoribonuclease XRN1, preferentially impacts transcripts with certain sequences or in particular functional categories.RNA. 2011 Mar;17(3):501-11. doi: 10.1261/rna.2467911. Epub 2011 Jan 11. RNA. 2011. PMID: 21224377 Free PMC article.
-
Monitoring of XRN4 Targets Reveals the Importance of Cotranslational Decay during Arabidopsis Development.Plant Physiol. 2020 Nov;184(3):1251-1262. doi: 10.1104/pp.20.00942. Epub 2020 Sep 10. Plant Physiol. 2020. PMID: 32913043 Free PMC article.
-
Arabidopsis mRNA decay landscape shaped by XRN 5'-3' exoribonucleases.Plant J. 2023 May;114(4):895-913. doi: 10.1111/tpj.16181. Epub 2023 Mar 28. Plant J. 2023. PMID: 36987558
-
Activity and roles of Arabidopsis thaliana XRN family exoribonucleases in noncoding RNA pathways.J Plant Res. 2017 Jan;130(1):25-31. doi: 10.1007/s10265-016-0887-z. Epub 2016 Dec 17. J Plant Res. 2017. PMID: 27988817 Review.
-
Mechanistic insights into DXO1 and XRN3: regulatory roles of RNA stability, transcription, and liquid-liquid phase separation in Arabidopsis thaliana (L.) Heynh.Plant Sci. 2025 Apr;353:112413. doi: 10.1016/j.plantsci.2025.112413. Epub 2025 Feb 3. Plant Sci. 2025. PMID: 39909287 Review.
Cited by
-
Metabolite Control of Translation by Conserved Peptide uORFs: The Ribosome as a Metabolite Multisensor.Plant Physiol. 2020 Jan;182(1):110-122. doi: 10.1104/pp.19.00940. Epub 2019 Aug 26. Plant Physiol. 2020. PMID: 31451550 Free PMC article.
-
TERA-Seq: true end-to-end sequencing of native RNA molecules for transcriptome characterization.Nucleic Acids Res. 2021 Nov 18;49(20):e115. doi: 10.1093/nar/gkab713. Nucleic Acids Res. 2021. PMID: 34428294 Free PMC article.
-
Rapid Recovery Gene Downregulation during Excess-Light Stress and Recovery in Arabidopsis.Plant Cell. 2017 Aug;29(8):1836-1863. doi: 10.1105/tpc.16.00828. Epub 2017 Jul 13. Plant Cell. 2017. PMID: 28705956 Free PMC article.
-
mRNA ADENOSINE METHYLASE promotes drought tolerance through N6-methyladenosine-dependent and independent impacts on mRNA regulation in Arabidopsis.New Phytol. 2025 Jan;245(1):183-199. doi: 10.1111/nph.20227. Epub 2024 Oct 27. New Phytol. 2025. PMID: 39462792 Free PMC article.
-
Structural and molecular mechanisms for the control of eukaryotic 5'-3' mRNA decay.Nat Struct Mol Biol. 2018 Dec;25(12):1077-1085. doi: 10.1038/s41594-018-0164-z. Epub 2018 Dec 5. Nat Struct Mol Biol. 2018. PMID: 30518847 Review.
References
-
- Chekanova J.A., et al. (2007). Genome-wide high-resolution mapping of exosome substrates reveals hidden features in the Arabidopsis transcriptome. Cell 131: 1340–1353. - PubMed
-
- Chiba Y., Green P. (2009). mRNA degradation machinery in plants. J. Plant Biol. 52: 114–124.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
