

HMGB1 Activates Proinflammatory Signaling via TLR5 Leading to Allodynia
- PMID: 27760316
- PMCID: PMC5087801
- DOI: 10.1016/j.celrep.2016.09.076
HMGB1 Activates Proinflammatory Signaling via TLR5 Leading to Allodynia
Abstract
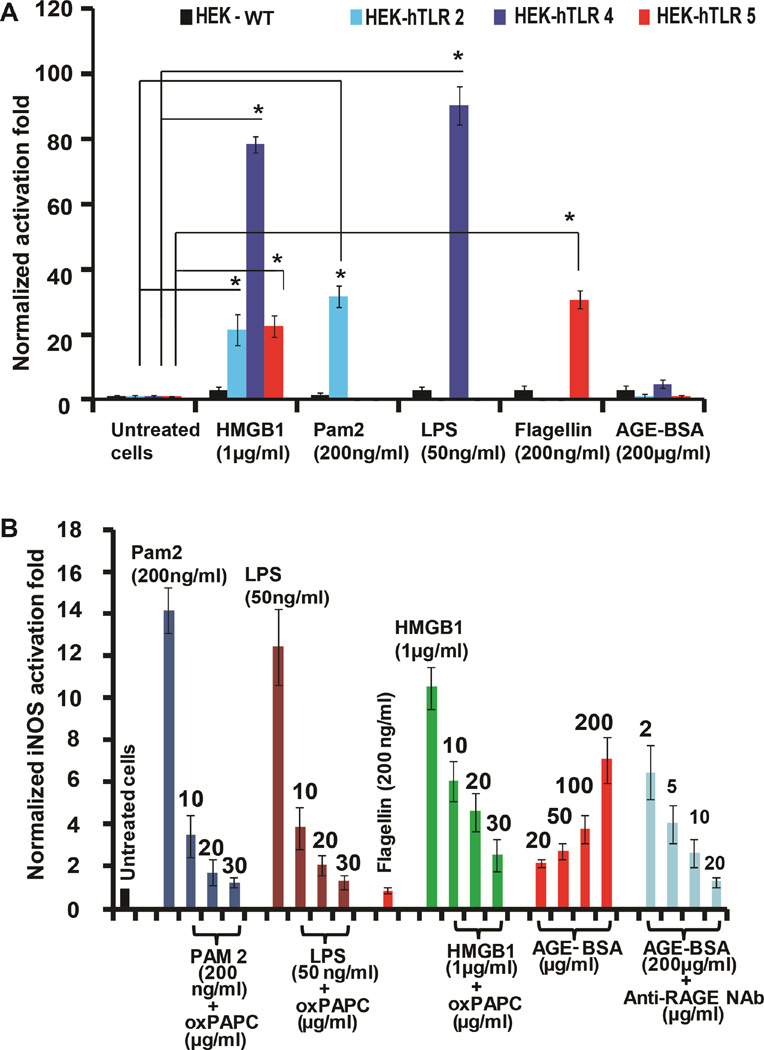
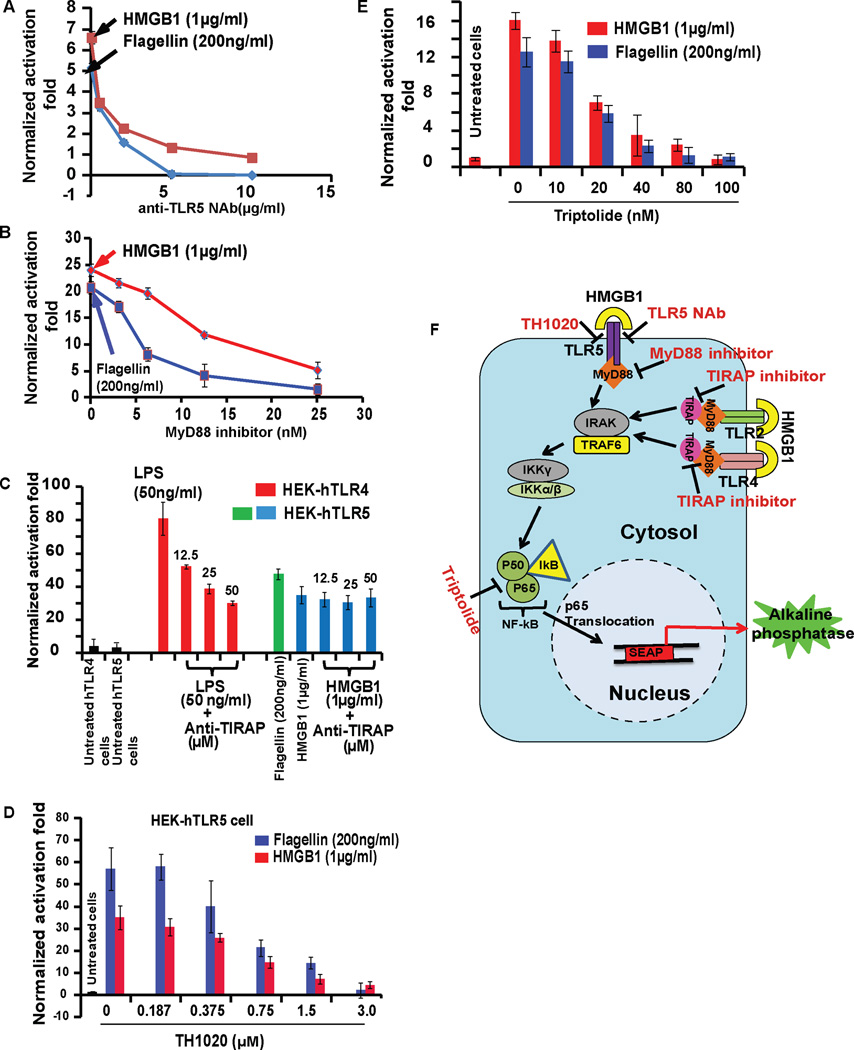
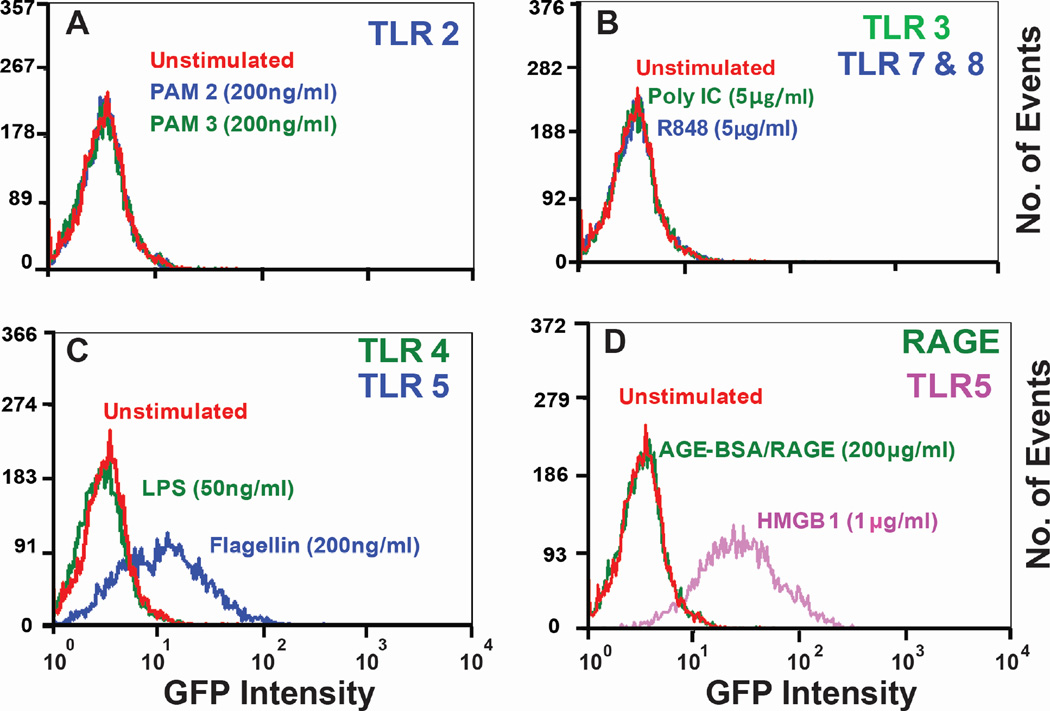
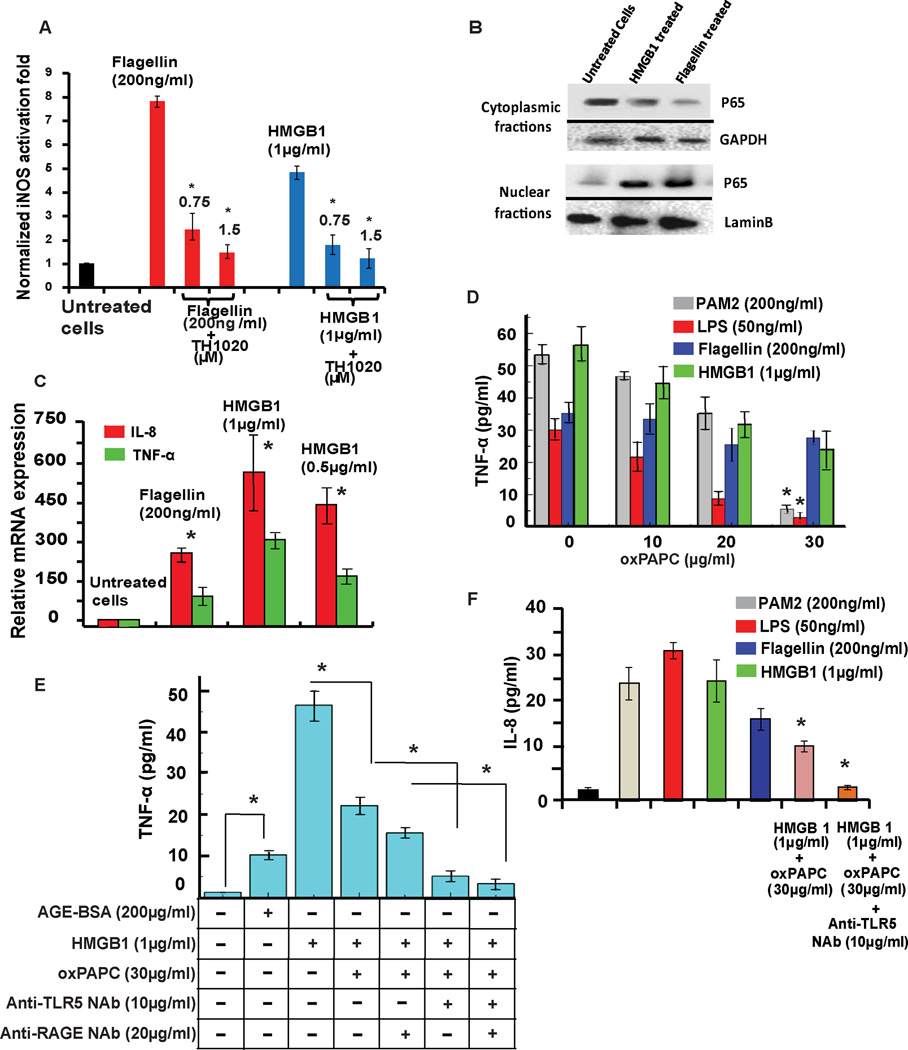
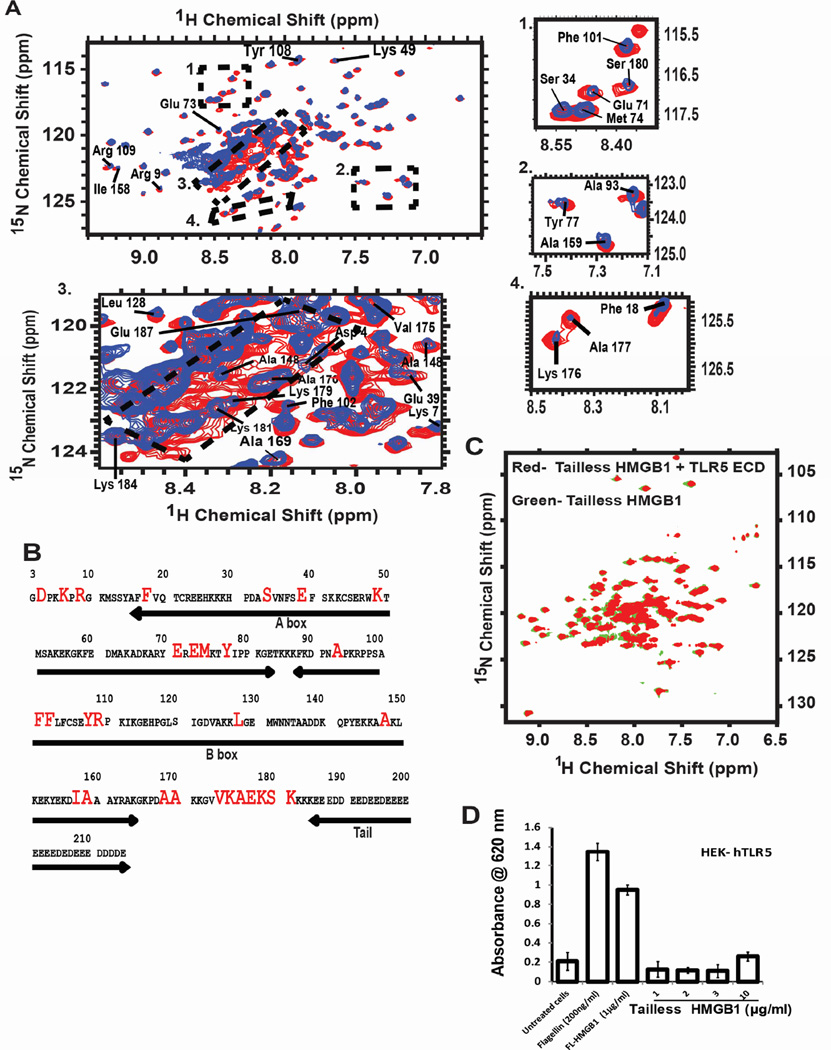
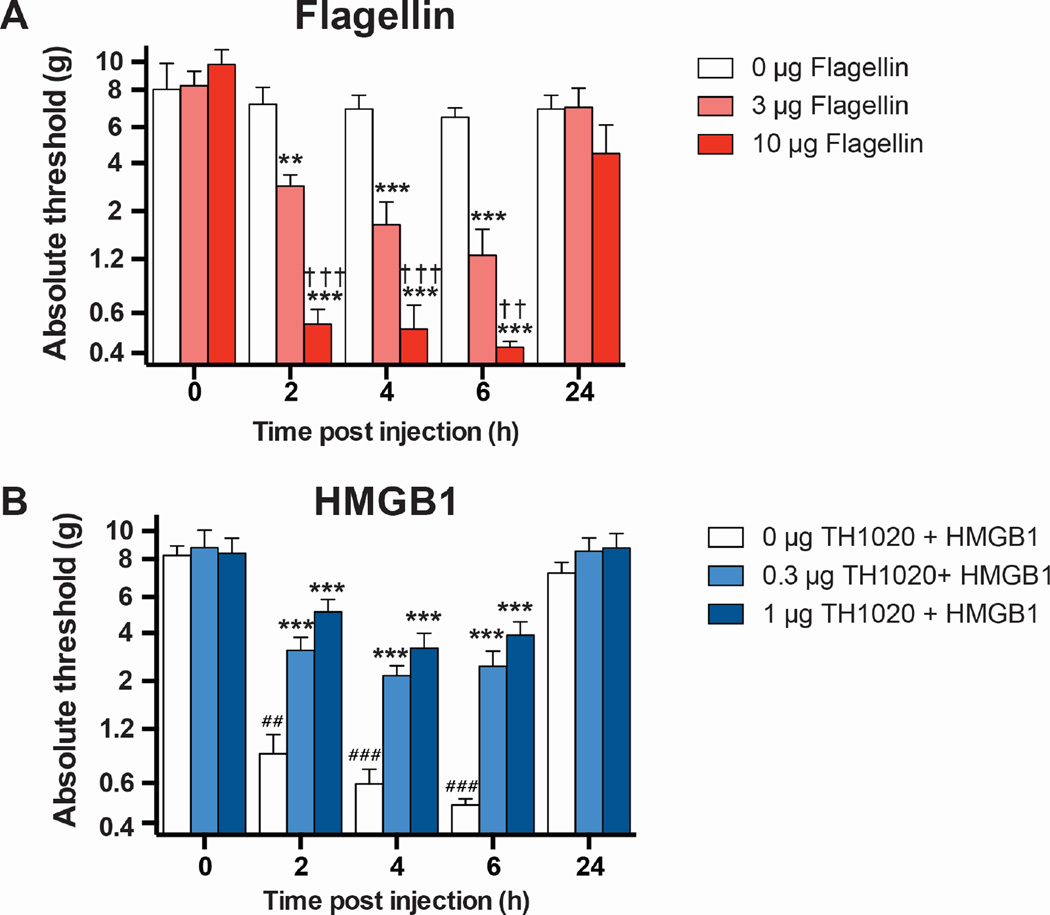
Infectious and sterile inflammatory diseases are correlated with increased levels of high mobility group box 1 (HMGB1) in tissues and serum. Extracellular HMGB1 is known to activate Toll-like receptors (TLRs) 2 and 4 and RAGE (receptor for advanced glycation endproducts) in inflammatory conditions. Here, we find that TLR5 is also an HMGB1 receptor that was previously overlooked due to lack of functional expression in the cell lines usually used for studying TLR signaling. HMGB1 binding to TLR5 initiates the activation of NF-κB signaling pathway in a MyD88-dependent manner, resulting in proinflammatory cytokine production and pain enhancement in vivo. Biophysical and in vitro results highlight an essential role for the C-terminal tail region of HMGB1 in facilitating interactions with TLR5. These results suggest that HMGB1-modulated TLR5 signaling is responsible for pain hypersensitivity.
Keywords: HMGB1; Toll-like receptors; cytokines; inflammation; pain.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. 2004;4:499–511. - PubMed
-
- Aravalli RN, Peterson PK, Lokensgard JR. Toll-like receptors in defense and damage of the central nervous system. J Neuroimmune Pharmacol. 2007;2:297–312. - PubMed
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
