

YUCCA auxin biosynthetic genes are required for Arabidopsis shade avoidance
- PMID: 27761349
- PMCID: PMC5068344
- DOI: 10.7717/peerj.2574
YUCCA auxin biosynthetic genes are required for Arabidopsis shade avoidance
Abstract
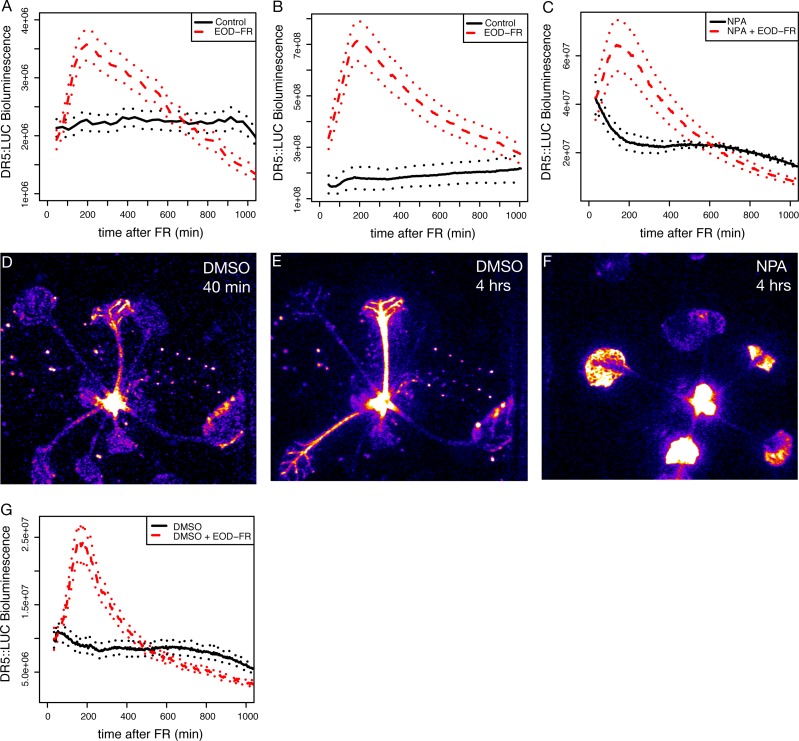
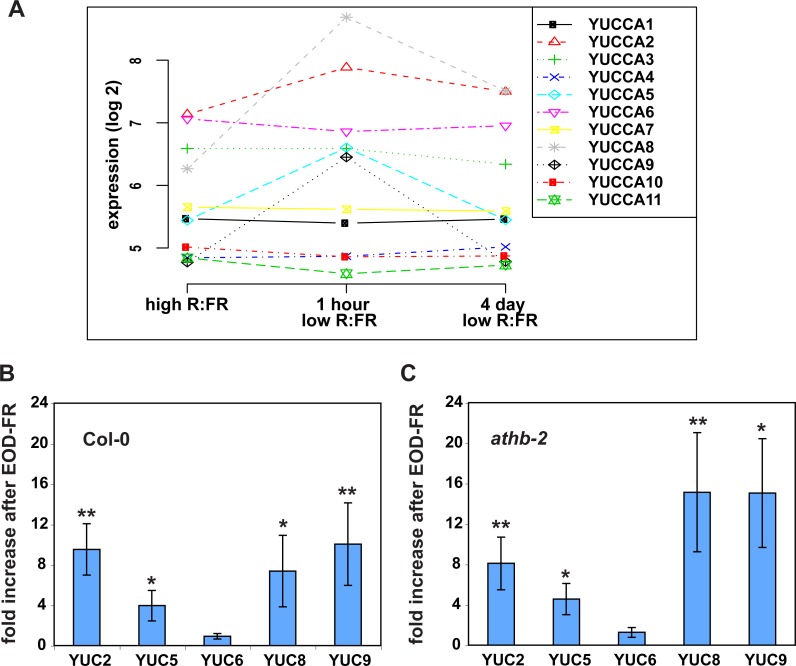
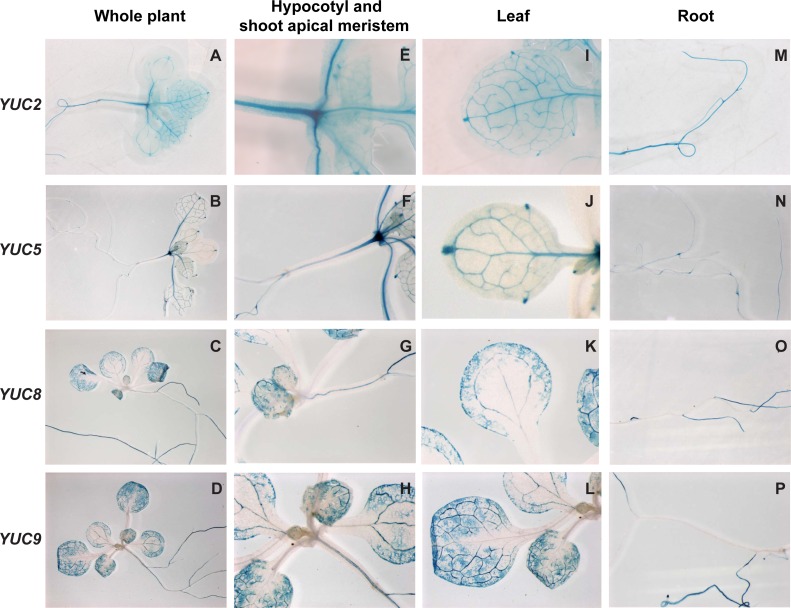
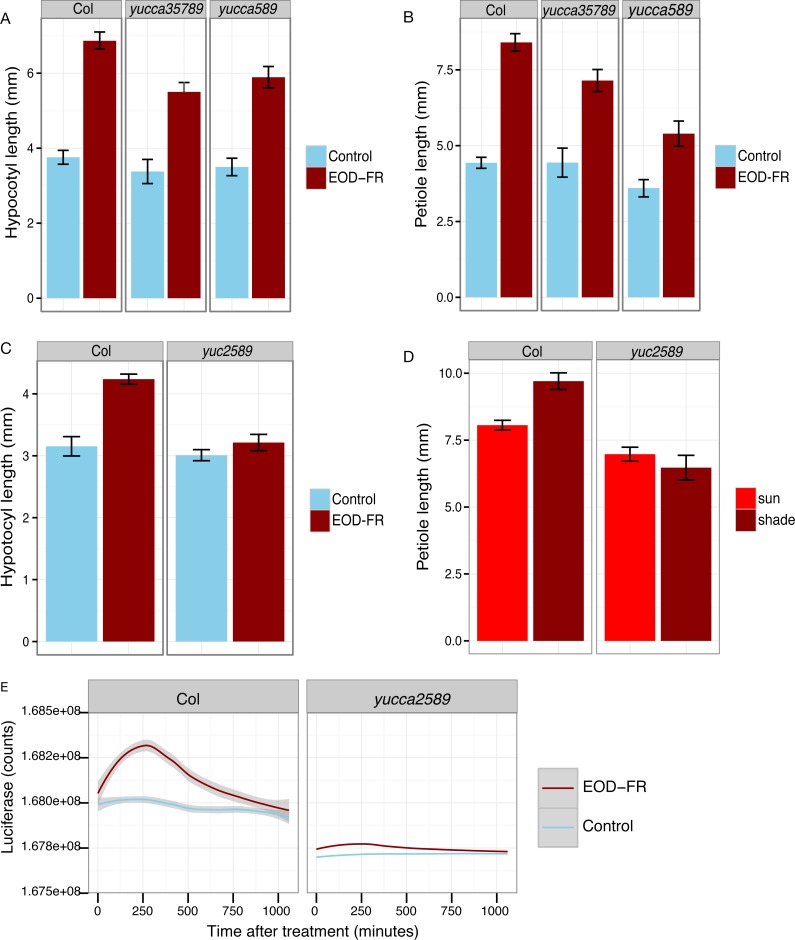
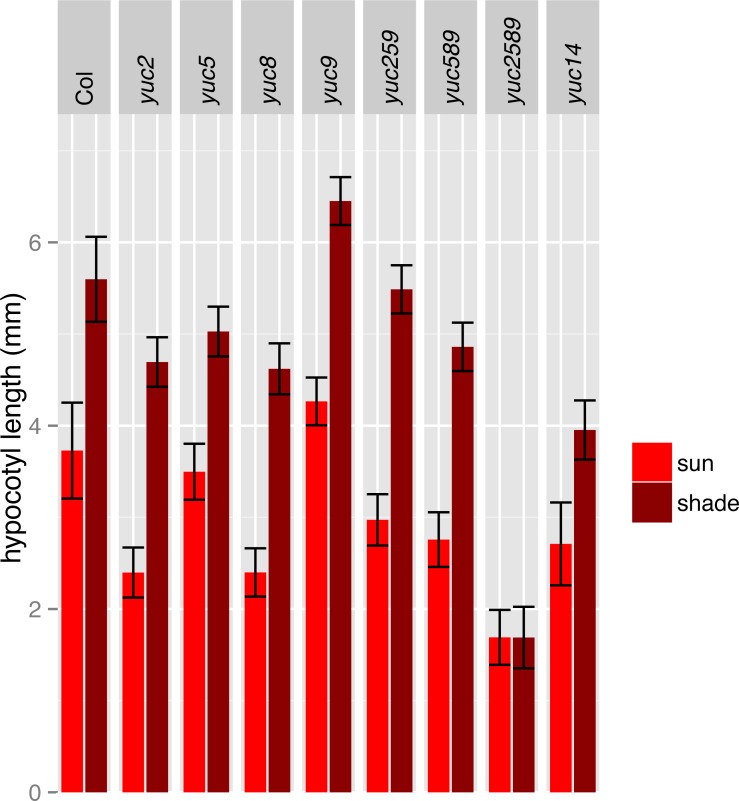
Plants respond to neighbor shade by increasing stem and petiole elongation. Shade, sensed by phytochrome photoreceptors, causes stabilization of PHYTOCHROME INTERACTING FACTOR proteins and subsequent induction of YUCCA auxin biosynthetic genes. To investigate the role of YUCCA genes in phytochrome-mediated elongation, we examined auxin signaling kinetics after an end-of-day far-red (EOD-FR) light treatment, and found that an auxin responsive reporter is rapidly induced within 2 hours of far-red exposure. YUCCA2, 5, 8, and 9 are all induced with similar kinetics suggesting that they could act redundantly to control shade-mediated elongation. To test this hypothesis we constructed a yucca2, 5, 8, 9 quadruple mutant and found that the hypocotyl and petiole EOD-FR and shade avoidance responses are completely disrupted. This work shows that YUCCA auxin biosynthetic genes are essential for detectable shade avoidance and that YUCCA genes are important for petiole shade avoidance.
Keywords: Auxin; Photomorphogenesis; Phytochrome; Shade avoidance.
Conflict of interest statement
Melissa Pytlak is an employee of PASCO scientific, Roseville, California and Andreah Wallace is an employee of Clontech Laboratories, Mountain View, California
Figures

References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, Gadrinab C, Heller C, Jeske A, Koesema E, Meyers CC, Parker H, Prednis L, Ansari Y, Choy N, Deen H, Geralt M, Hazari N, Hom E, Karnes M, Mulholland C, Ndubaku R, Schmidt I, Guzman P, Aguilar-Henonin L, Schmid M, Weigel D, Carter DE, Marchand T, Risseeuw E, Brogden D, Zeko A, Crosby WL, Berry CC, Ecker JR. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science. 2003;301:653–657. doi: 10.1126/science.1086391. - DOI - PubMed
-
- Bates D, Mächler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4. Journal of Statistical Software. 2015;67(1):1–48.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
