

Specific and essential but not sufficient roles of LRRC8A in the activity of volume-sensitive outwardly rectifying anion channel (VSOR)
- PMID: 27764579
- PMCID: PMC5398601
- DOI: 10.1080/19336950.2016.1247133
Specific and essential but not sufficient roles of LRRC8A in the activity of volume-sensitive outwardly rectifying anion channel (VSOR)
Abstract
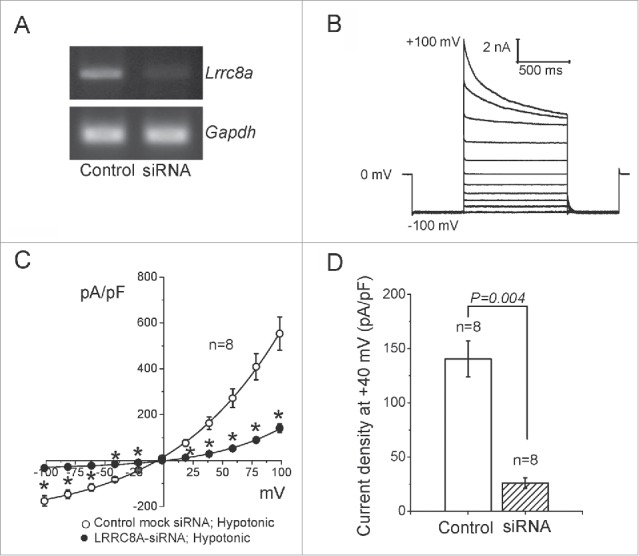
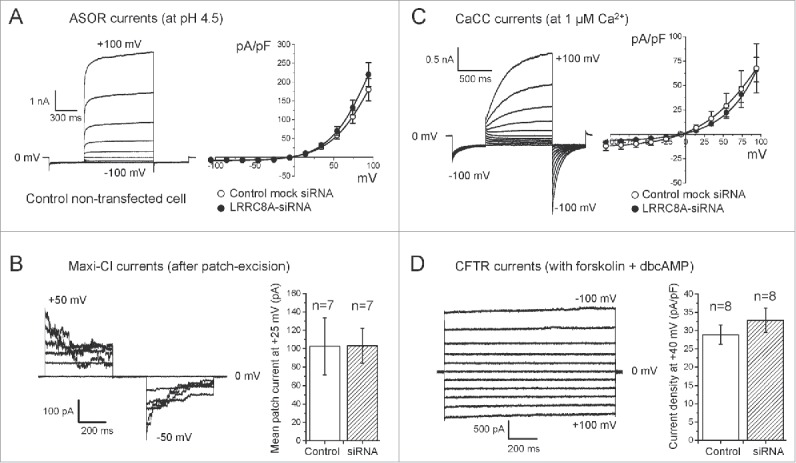
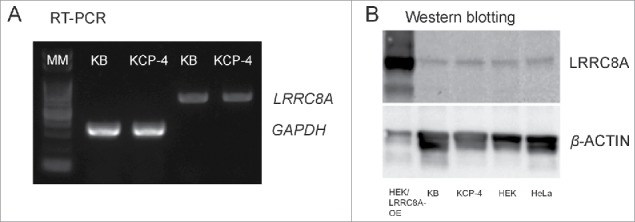
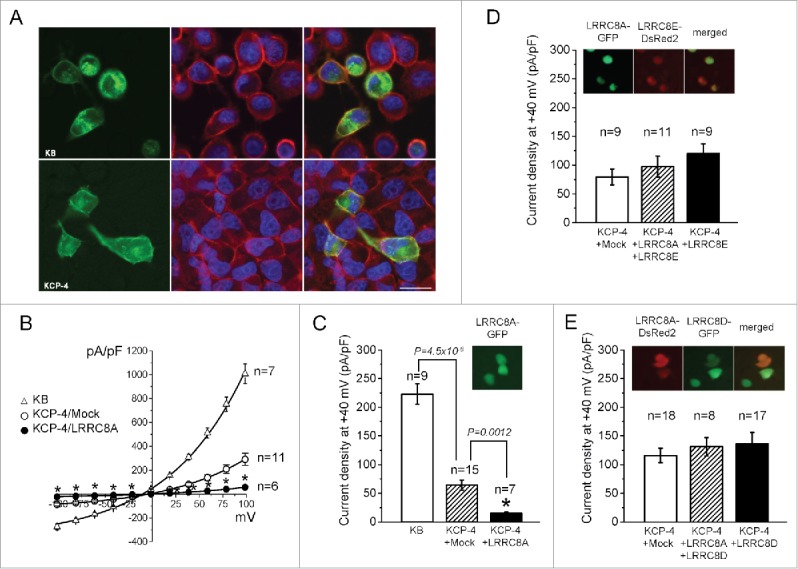
The broadly expressed volume-sensitive outwardly rectifying anion channel (VSOR, also called VRAC) plays essential roles in cell survival and death. Recent findings have suggested that LRRC8A is a core component of VSOR in human cells. In the present study, VSOR currents were found to be largely reduced by siRNA against LRRC8A in mouse C127 cells as well. In contrast, LRRC8A knockdown never affected activities of 4 other types of anion channel activated by acid, Ca2+, patch excision or cAMP. While cisplatin-resistant KCP-4 cells poorly expressed endogenous VSOR activity, molecular expression levels of LRRC8A, LRRC8D and LRRC8E were indistinguishable between VSOR-deficient KCP-4 cells and the parental VSOR-rich KB cells. Furthermore, overexpression of LRRC8A alone or together with LRRC8D or LRRC8E in KCP-4 cells failed to restore VSOR activity. These results show that deficiency of VSOR currents in KCP-4 cells is not due to insufficient expression of the LRRC8A/D/E gene, suggesting an essential involvement of some other factor(s), and indicate that further study is required to better understand the complexities of the molecular determinants of VSOR, including the precise role of LRRC8 proteins.
Keywords: LRRC8; VSOR; anion channel; cisplatin resistance; volume-sensitive.
Figures
Similar articles
-
Distinct contributions of LRRC8A and its paralogs to the VSOR anion channel from those of the ASOR anion channel.Channels (Austin). 2017 Mar 4;11(2):167-172. doi: 10.1080/19336950.2016.1230574. Epub 2016 Aug 31. Channels (Austin). 2017. PMID: 27579940 Free PMC article.
-
Distinct pharmacological and molecular properties of the acid-sensitive outwardly rectifying (ASOR) anion channel from those of the volume-sensitive outwardly rectifying (VSOR) anion channel.Pflugers Arch. 2016 May;468(5):795-803. doi: 10.1007/s00424-015-1786-1. Epub 2016 Jan 8. Pflugers Arch. 2016. PMID: 26743872
-
Molecular composition and heterogeneity of the LRRC8-containing swelling-activated osmolyte channels in primary rat astrocytes.J Physiol. 2017 Nov 15;595(22):6939-6951. doi: 10.1113/JP275053. Epub 2017 Sep 12. J Physiol. 2017. PMID: 28833202 Free PMC article.
-
Biophysics and Physiology of the Volume-Regulated Anion Channel (VRAC)/Volume-Sensitive Outwardly Rectifying Anion Channel (VSOR).Pflugers Arch. 2016 Mar;468(3):371-83. doi: 10.1007/s00424-015-1781-6. Epub 2016 Jan 6. Pflugers Arch. 2016. PMID: 26739710 Review.
-
VRAC: molecular identification as LRRC8 heteromers with differential functions.Pflugers Arch. 2016 Mar;468(3):385-93. doi: 10.1007/s00424-015-1766-5. Epub 2015 Dec 3. Pflugers Arch. 2016. PMID: 26635246 Review.
Cited by
-
A FRET sensor of C-terminal movement reveals VRAC activation by plasma membrane DAG signaling rather than ionic strength.Elife. 2019 Jun 18;8:e45421. doi: 10.7554/eLife.45421. Elife. 2019. PMID: 31210638 Free PMC article.
-
Sphingosine-1-Phosphate Induces ATP Release via Volume-Regulated Anion Channels in Breast Cell Lines.Life (Basel). 2021 Aug 19;11(8):851. doi: 10.3390/life11080851. Life (Basel). 2021. PMID: 34440595 Free PMC article.
-
Biophysics and Structure-Function Relationships of LRRC8-Formed Volume-Regulated Anion Channels.Biophys J. 2019 Apr 2;116(7):1185-1193. doi: 10.1016/j.bpj.2019.02.014. Epub 2019 Feb 26. Biophys J. 2019. PMID: 30871717 Free PMC article. Review.
-
Physiology of the volume-sensitive/regulatory anion channel VSOR/VRAC. Part 1: from its discovery and phenotype characterization to the molecular entity identification.J Physiol Sci. 2024 Jan 18;74(1):3. doi: 10.1186/s12576-023-00897-x. J Physiol Sci. 2024. PMID: 38238667 Free PMC article. Review.
-
Tweety-homolog (Ttyh) Family Encodes the Pore-forming Subunits of the Swelling-dependent Volume-regulated Anion Channel (VRACswell) in the Brain.Exp Neurobiol. 2019 Apr;28(2):183-215. doi: 10.5607/en.2019.28.2.183. Epub 2019 Apr 30. Exp Neurobiol. 2019. PMID: 31138989 Free PMC article.
References
-
- Okada Y, Sato K, Toychiev AH, Suzuki M, Dutta AK, Inoue H, Sabirov RZ. The puzzles of volume-activated anion channels In Physiology and Pathology of Chloride Transporters and Channels in the Nervous System. From Molecules to Diseases, Alvarez-Leefmans FJ, Delpire E, eds. (San Diego: Elsevier; ) 2009; pp. 283-306
-
- Pedersen SF, Okada Y, Nilius B. Biophysics and physiology of the volume-regulated anion channel (VRAC)/volume-sensitive outwardly rectifying anion channel (VSOR). Pflugers Arch Eur J Physiol 2016; 468:371-83; http://dx.doi.org/10.1007/s00424-015-1781-6 - DOI - PubMed
-
- Valverde MA, Diaz M, Sepúlveda FV, Gill DR, Hyde SC, Higgins CF. Volume-regulated chloride channels associated with the human multidrug-resistance P-glycoprotein. Nature 1992; 355:830-3; PMID:1371598; http://dx.doi.org/10.1038/355830a0 - DOI - PubMed
-
- Paulmichl M, Li Y, Wickman K, Ackerman M, Peralta E, Clapham D. New mammalian chloride channel identified by expression cloning. Nature 1992; 356:238-41; PMID:1313151; http://dx.doi.org/10.1038/356238a0 - DOI - PubMed
-
- Duan D, Winter C, Cowley S, Hume JR, Horowitz B. Molecular identification of a volume-regulated chloride channel. Nature 1997; 390:417-21; PMID:9389484; http://dx.doi.org/10.1038/37151 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous