

A combined gene expression and functional study reveals the crosstalk between N-Myc and differentiation-inducing microRNAs in neuroblastoma cells
- PMID: 27764804
- PMCID: PMC5346721
- DOI: 10.18632/oncotarget.12676
A combined gene expression and functional study reveals the crosstalk between N-Myc and differentiation-inducing microRNAs in neuroblastoma cells
Abstract
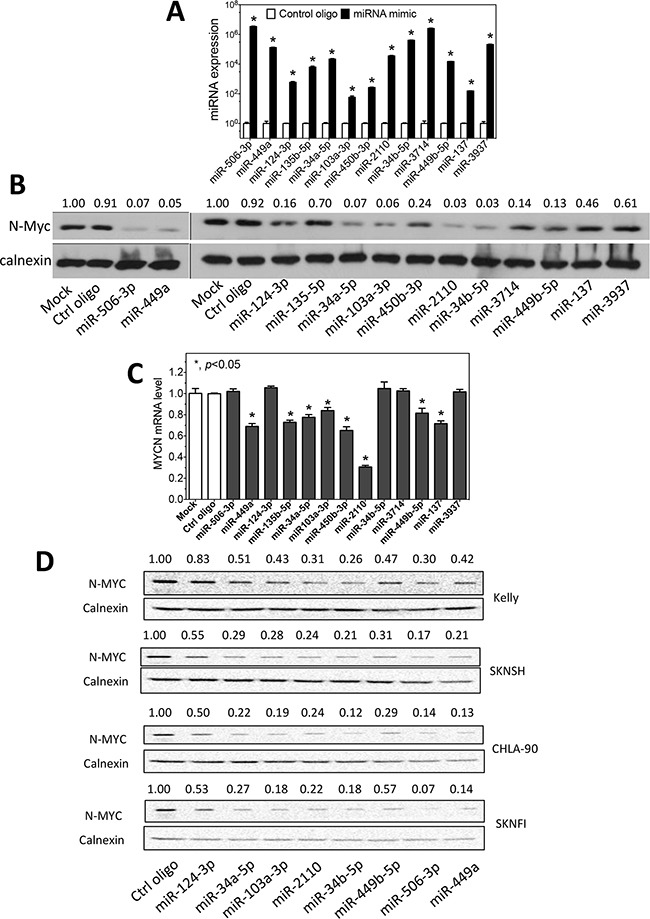
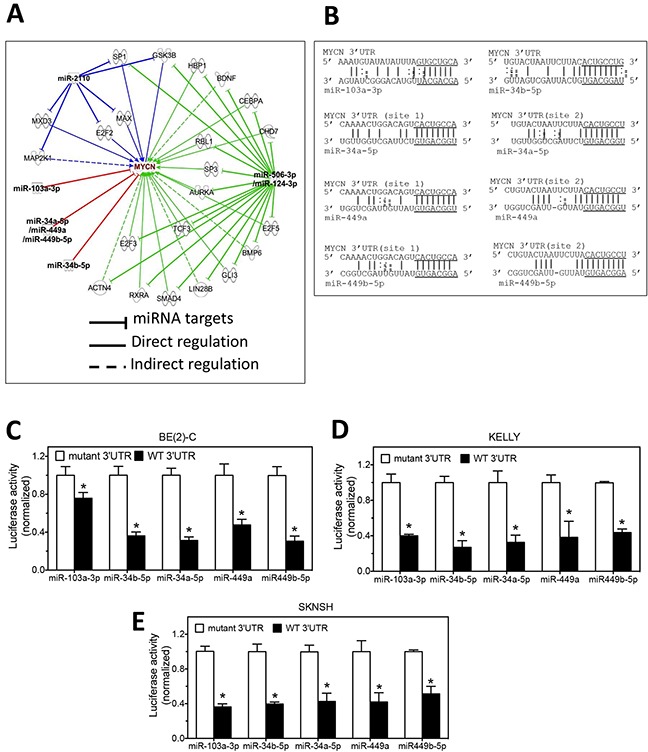
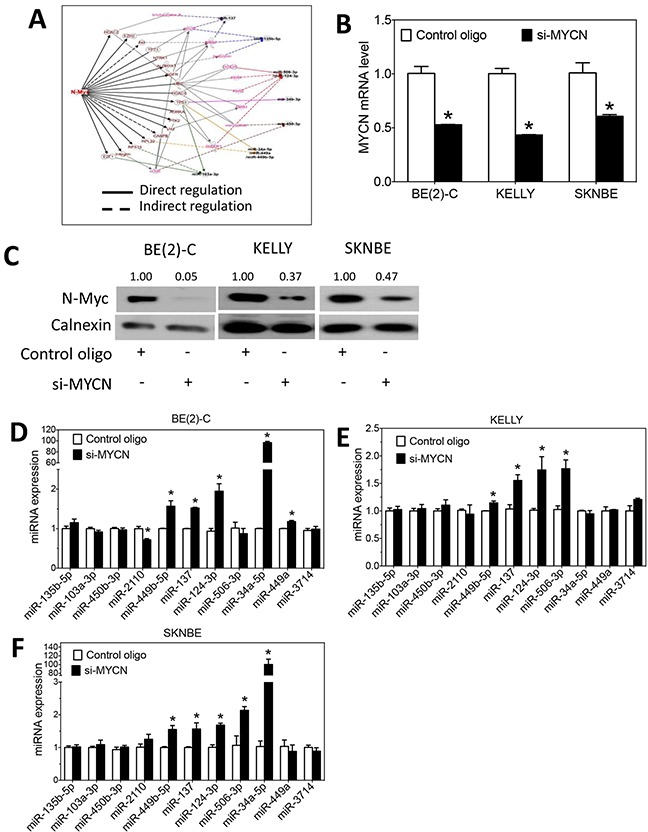
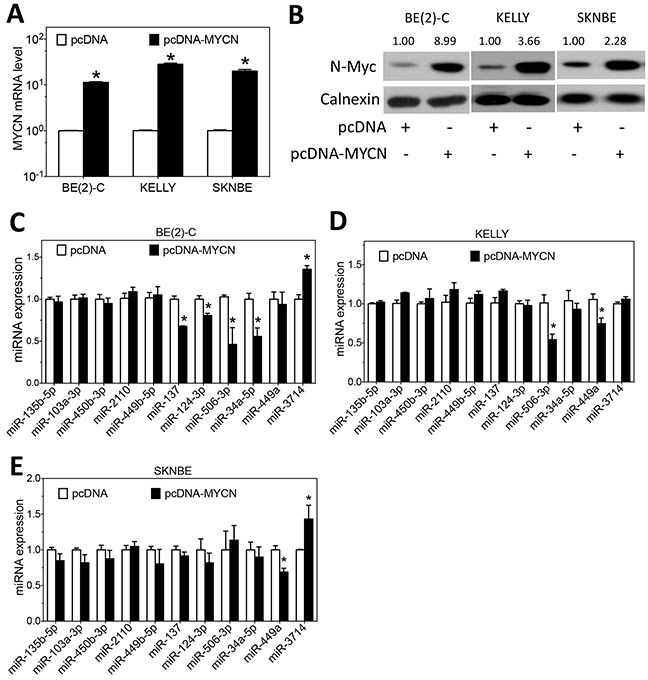
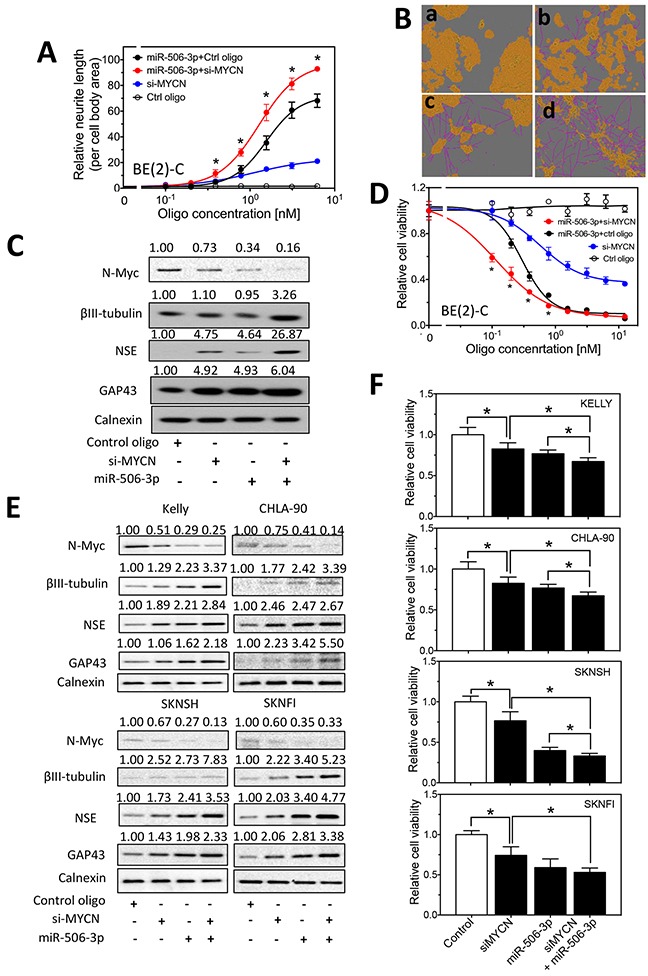
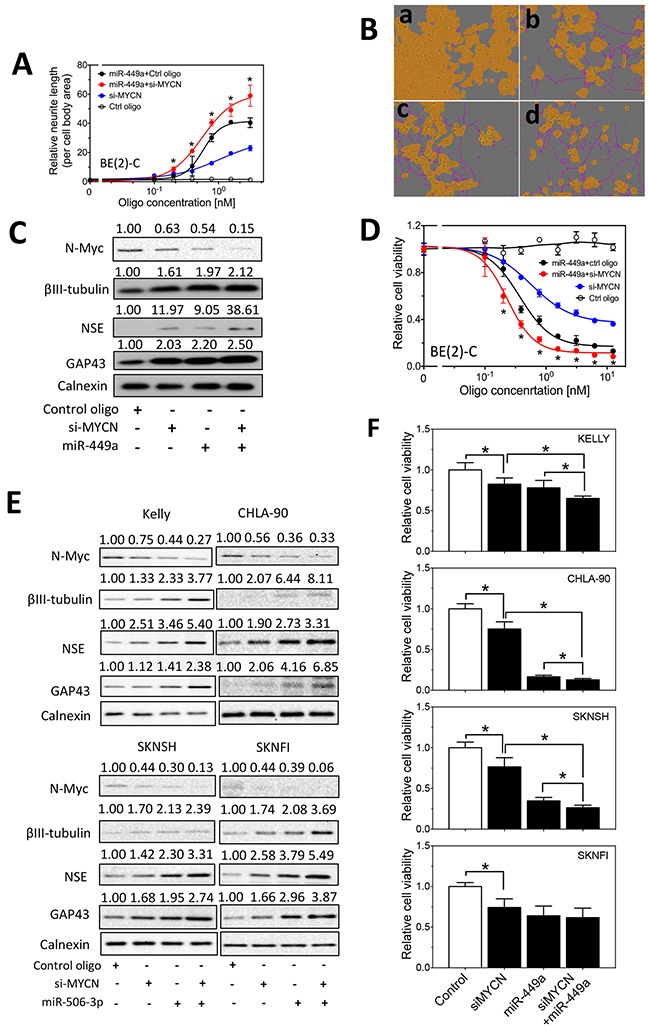
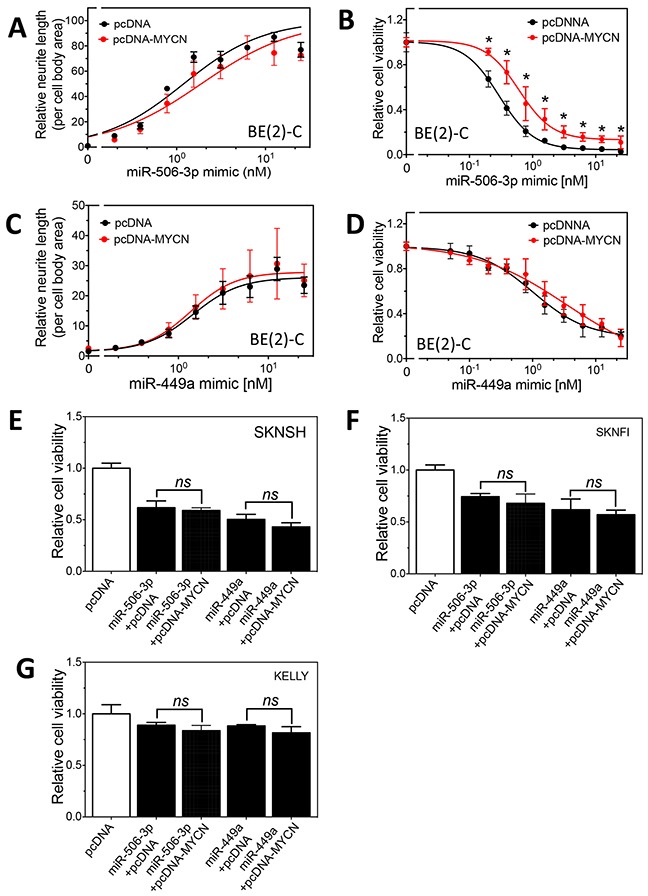
MYCN amplification is the most common genetic alteration in neuroblastoma and plays a critical role in neuroblastoma tumorigenesis. MYCN regulates neuroblastoma cell differentiation, which is one of the mechanisms underlying its oncogenic function. We recently identified a group of differentiation-inducing microRNAs. Given the demonstrated inter-regulation between MYCN and microRNAs, we speculated that MYCN and the differentiation-inducing microRNAs might form an interaction network to control the differentiation of neuroblastoma cells. In this study, we found that eight of the thirteen differentiation-inducing microRNAs, miR-506-3p, miR-124-3p, miR-449a, miR-34a-5p, miR-449b-5p, miR-103a-3p, miR-2110 and miR-34b-5p, inhibit N-Myc expression by either directly targeting the MYCN 3'UTR or through indirect regulations. Further investigation showed that both MYCN-dependent and MYCN-independent pathways play roles in mediating the differentiation-inducing function of miR-506-3p and miR-449a, two microRNAs that dramatically down-regulate MYCN expression. On the other hand, we found that N-Myc inhibits the expression of multiple differentiation-inducing microRNAs, suggesting that these miRNAs play a role in mediating the function of MYCN. In examining the published dataset collected from clinical neuroblastoma specimens, we found that expressions of two miRNAs, miR-137 and miR-2110, were significantly anti-correlated with MYCN mRNA levels, suggesting their interactions with MYCN play a clinically-relevant role in maintaining the MYCN and miRNA expression levels in neuroblastoma. Our findings altogether suggest that MYCN and differentiation-inducing miRNAs form an interaction network that play an important role in neuroblastoma tumorigenesis through regulating cell differentiation.
Keywords: MYCN; differentiation; microRNA; neuroblastoma.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Brodeur GM, Seeger RC, Schwab M, Varmus HE, Bishop JM. Amplification of N-myc in untreated human neuroblastomas correlates with advanced disease stage. Science. 1984;224:1121–1124. - PubMed
-
- Brodeur GM. Neuroblastoma: biological insights into a clinical enigma. Nature reviews Cancer. 2003;3:203–216. - PubMed
-
- Seeger RC, Brodeur GM, Sather H, Dalton A, Siegel SE, Wong KY, Hammond D. Association of multiple copies of the N-myc oncogene with rapid progression of neuroblastomas. N Engl J Med. 1985;313:1111–1116. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
