

O6-2'-Deoxyguanosine-butylene-O6-2'-deoxyguanosine DNA Interstrand Cross-Links Are Replication-Blocking and Mutagenic DNA Lesions
- PMID: 27768841
- PMCID: PMC5665164
- DOI: 10.1021/acs.chemrestox.6b00278
O6-2'-Deoxyguanosine-butylene-O6-2'-deoxyguanosine DNA Interstrand Cross-Links Are Replication-Blocking and Mutagenic DNA Lesions
Abstract
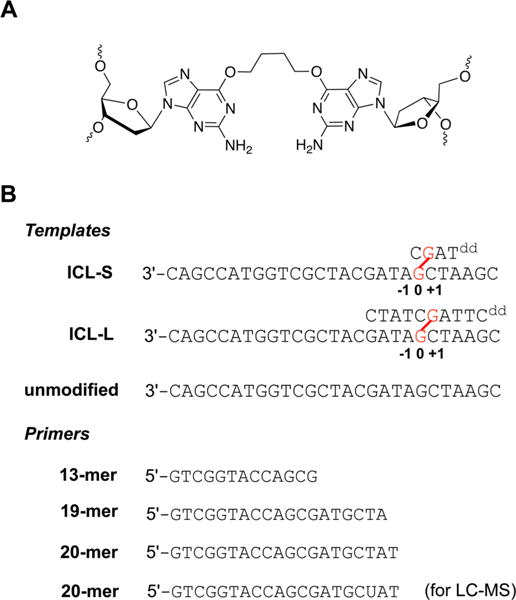
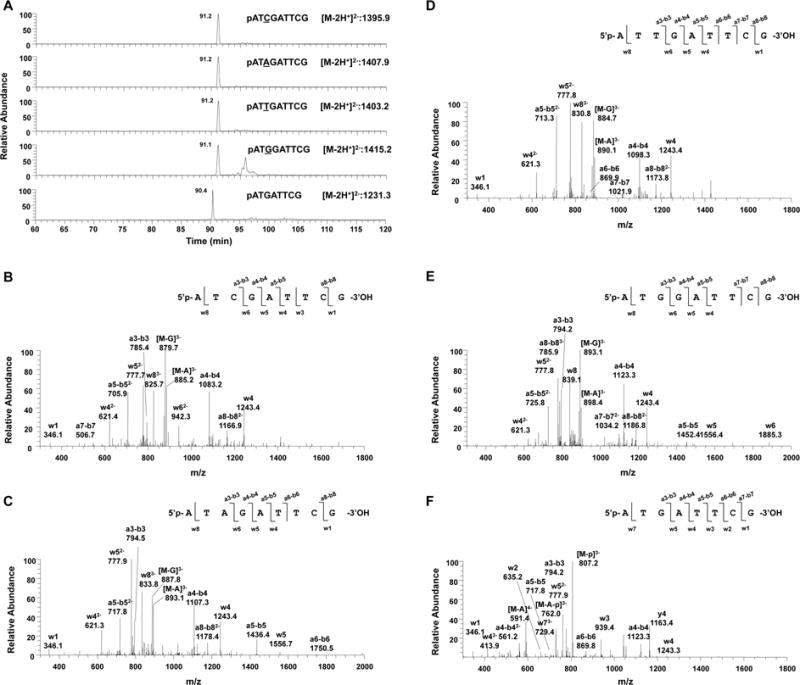
DNA interstrand cross-links (ICLs) are cytotoxic DNA lesions derived from reactions of DNA with a number of anti-cancer reagents as well as endogenous bifunctional electrophiles. Deciphering the DNA repair mechanisms of ICLs is important for understanding the toxicity of DNA cross-linking agents and for developing effective chemotherapies. Previous research has focused on ICLs cross-linked with the N7 and N2 atoms of guanine as well as those formed at the N6 atom of adenine; however, little is known about the mutagenicity of O6-dG-derived ICLs. Although less abundant, O6-alkylated guanine DNA lesions are chemically stable and highly mutagenic. Here, O6-2'-deoxyguanosine-butylene-O6-2'-deoxyguanosine (O6-dG-C4-O6-dG) is designed as a chemically stable ICL, which can be induced by the action of bifunctional alkylating agents. We investigate the DNA replication-blocking and mutagenic properties of O6-dG-C4-O6-dG ICLs during an important step in ICL repair, translesion DNA synthesis (TLS). The model replicative DNA polymerase (pol) Sulfolobus solfataricus P2 DNA polymerase B1 (Dpo1) is able to incorporate a correct nucleotide opposite the cross-linked template guanine of ICLs with low efficiency and fidelity but cannot extend beyond the ICLs. Translesion synthesis by human pol κ is completely inhibited by O6-dG-C4-O6-dG ICLs. Moderate bypass activities are observed for human pol η and S. solfataricus P2 DNA polymerase IV (Dpo4). Among the pols tested, pol η exhibits the highest bypass activity; however, 70% of the bypass products are mutagenic containing substitutions or deletions. The increase in the size of unhooked repair intermediates elevates the frequency of deletion mutation. Lastly, the importance of pol η in O6-dG-derived ICL bypass is demonstrated using whole cell extracts of Xeroderma pigmentosum variant patient cells and those complemented with pol η. Together, this study provides the first set of biochemical evidence for the mutagenicity of O6-dG-derived ICLs.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





References
-
- Hübscher U, Maga G. DNA replication and repair bypass machines. Curr Opin Chem Biol. 2011;15:627–635. - PubMed
-
- Hoeijmakers JHJ. Genome maintenance mechanisms for preventing cancer. Nature. 2001;411:366–374. - PubMed
-
- Aguilera A, García-Muse T. Causes of genome instability. Annu Rev Genet. 2013;47:1–32. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
