

Chromatin Association of Gcn4 Is Limited by Post-translational Modifications Triggered by its DNA-Binding in Saccharomyces cerevisiae
- PMID: 27770033
- PMCID: PMC5161277
- DOI: 10.1534/genetics.116.194134
Chromatin Association of Gcn4 Is Limited by Post-translational Modifications Triggered by its DNA-Binding in Saccharomyces cerevisiae
Abstract
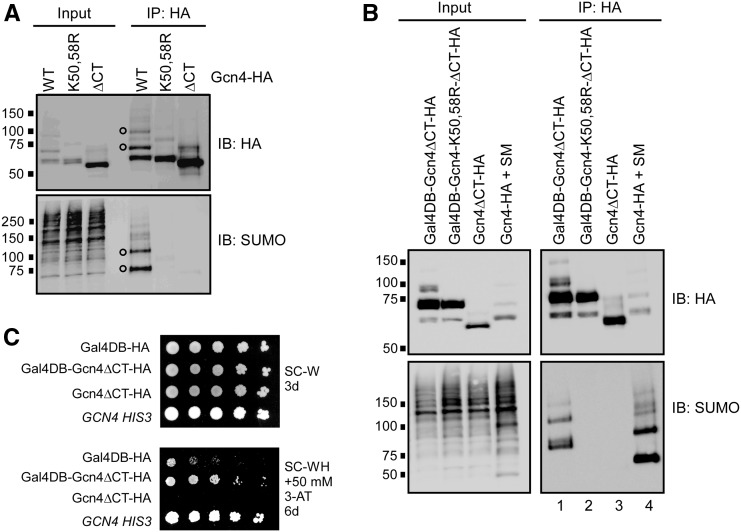
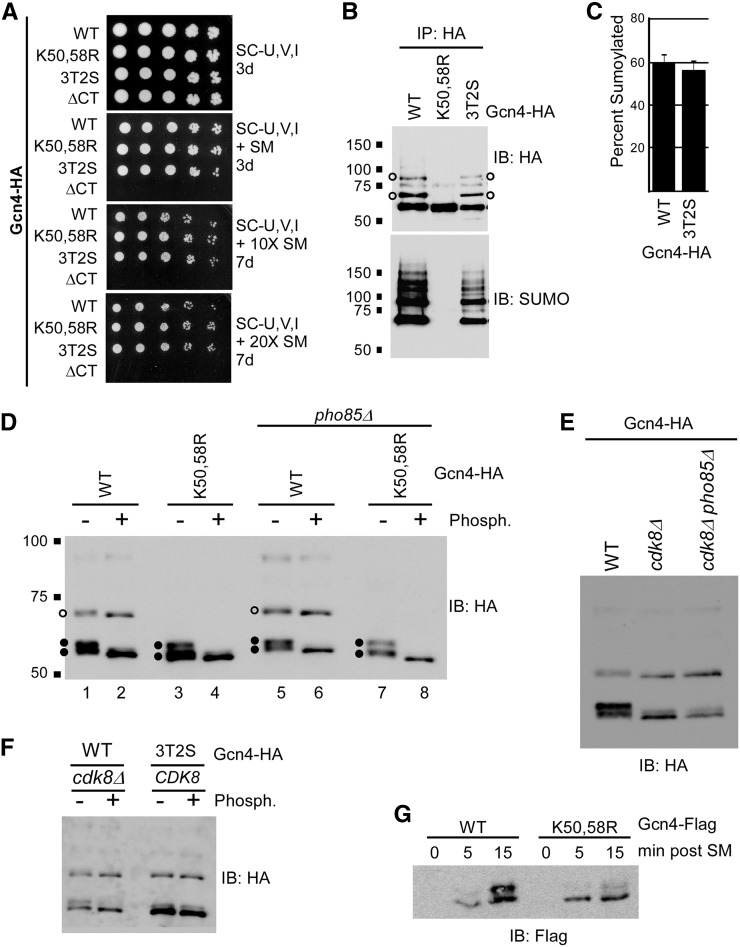
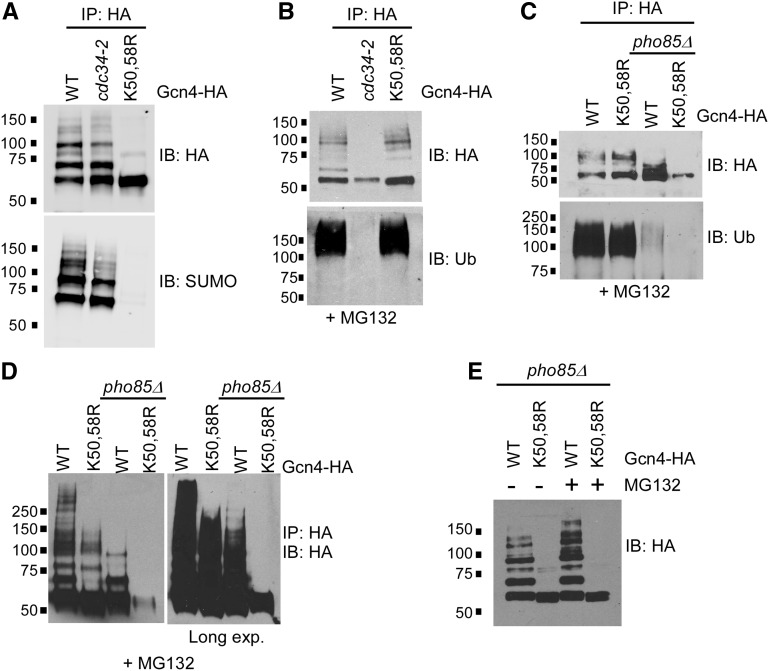
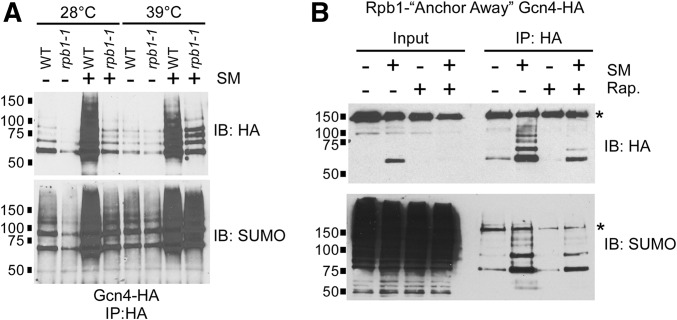
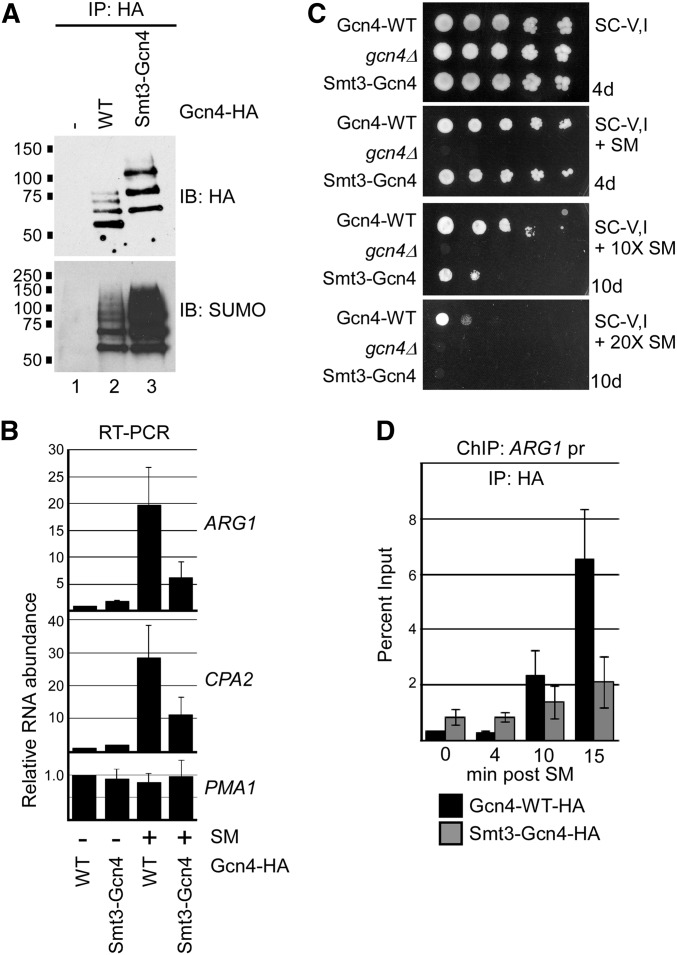
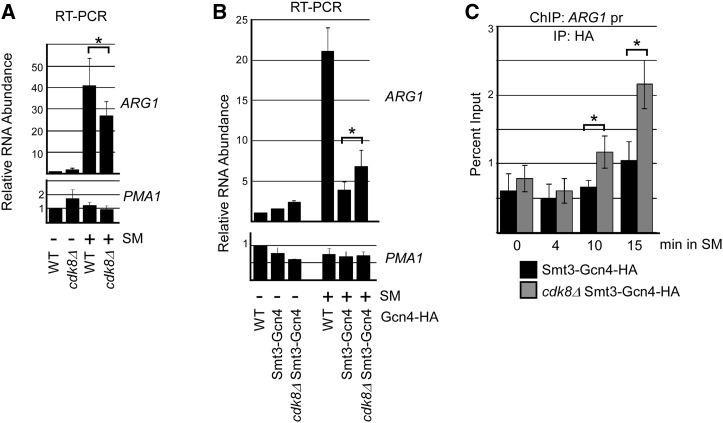
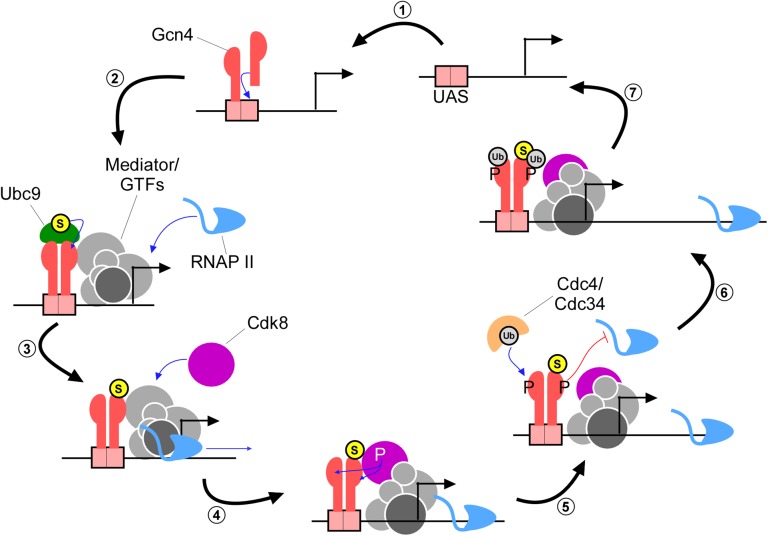
The Saccharomyces cerevisiae transcription factor Gcn4 is expressed during amino acid starvation, and its abundance is controlled by ubiquitin-mediated proteolysis. Cdk8, a kinase component of the RNA polymerase II Mediator complex, phosphorylates Gcn4, which triggers its ubiquitination/proteolysis, and is thought to link Gcn4 degradation with transcription of target genes. In addition to phosphorylation and ubiquitination, we previously showed that Gcn4 becomes sumoylated in a DNA-binding dependent manner, while a nonsumoylatable form of Gcn4 showed increased chromatin occupancy, but only if Cdk8 was present. To further investigate how the association of Gcn4 with chromatin is regulated, here we examine determinants for Gcn4 sumoylation, and how its post-translational modifications are coordinated. Remarkably, artificially targeting Gcn4 that lacks its DNA binding domain to a heterologous DNA site restores sumoylation at its natural modification sites, indicating that DNA binding is sufficient for the modification to occur in vivo Indeed, we find that neither transcription of target genes nor phosphorylation are required for Gcn4 sumoylation, but blocking its sumoylation alters its phosphorylation and ubiquitination patterns, placing Gcn4 sumoylation upstream of these Cdk8-mediated modifications. Strongly supporting a role for sumoylation in limiting its association with chromatin, a hyper-sumoylated form of Gcn4 shows dramatically reduced DNA occupancy and expression of target genes. Importantly, we find that Cdk8 is at least partly responsible for clearing hyper-sumoylated Gcn4 from DNA, further implicating sumoylation as a stimulus for Cdk8-mediated phosphorylation and degradation. These results support a novel function for SUMO in marking the DNA-bound form of a transcription factor, which triggers downstream processes that limit its association with chromatin, thus preventing uncontrolled expression of target genes.
Keywords: Cdk8; Gcn4; gene activation; sumoylation; transcription.
Copyright © 2016 by the Genetics Society of America.
Figures
References
-
- Amberg D. C., D. J. Burke, and J. N. Strathern, 2006 Yeast RNA isolation: small-scale. CSH Protoc. 2006: pii: pdb.prot4155. - PubMed
-
- Andrau J.-C., van de Pasch L., Lijnzaad P., Bijma T., Koerkamp M. G., et al. , 2006. Genome-wide location of the coactivator mediator: binding without activation and transient Cdk8 interaction on DNA. Mol. Cell 22: 179–192. - PubMed
-
- Bertolotto C., Lesueur F., Giuliano S., Strub T., de Lichy M., et al. , 2011. A SUMOylation-defective MITF germline mutation predisposes to melanoma and renal carcinoma. Nature 480: 94–98. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
