

Comprehensive proteome profiling of glioblastoma-derived extracellular vesicles identifies markers for more aggressive disease
- PMID: 27770278
- PMCID: PMC5306193
- DOI: 10.1007/s11060-016-2298-3
Comprehensive proteome profiling of glioblastoma-derived extracellular vesicles identifies markers for more aggressive disease
Abstract
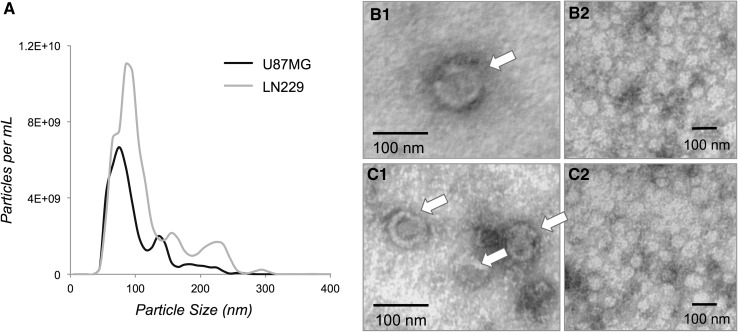
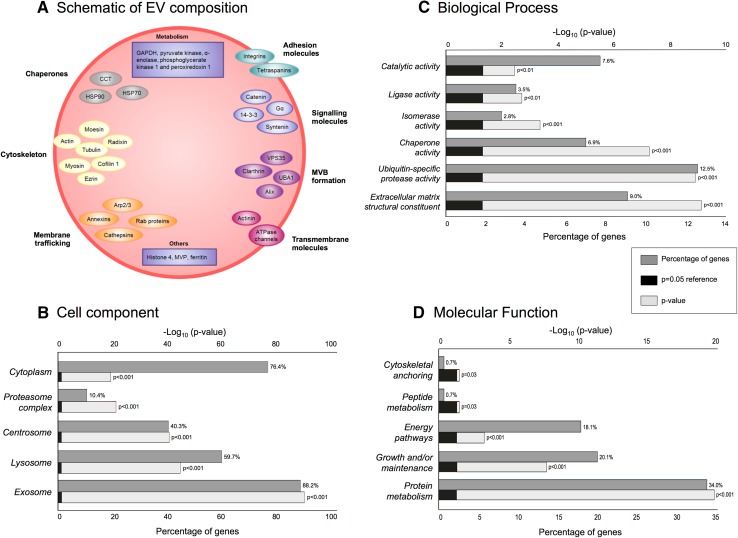
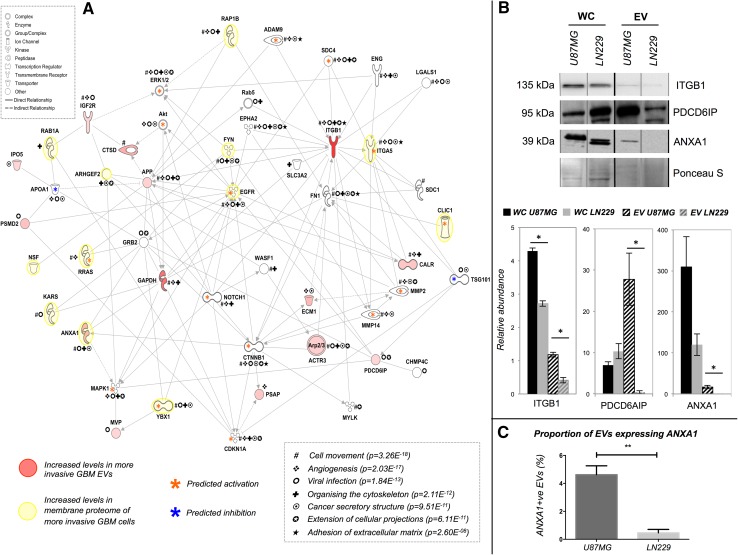
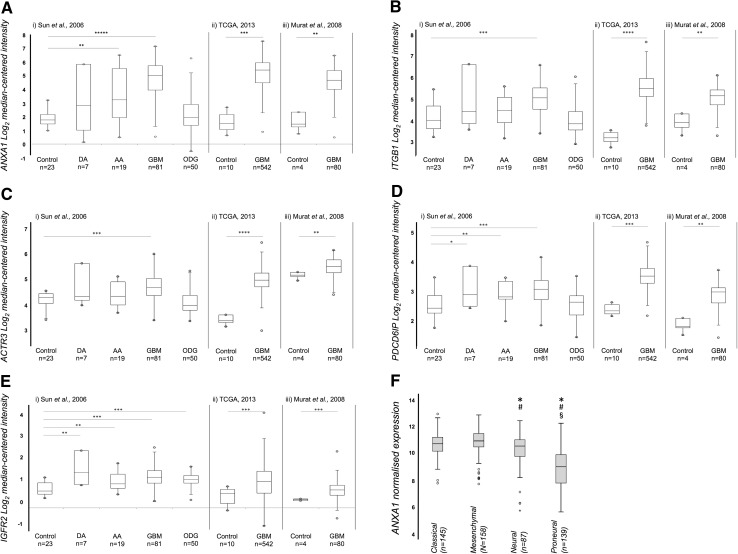
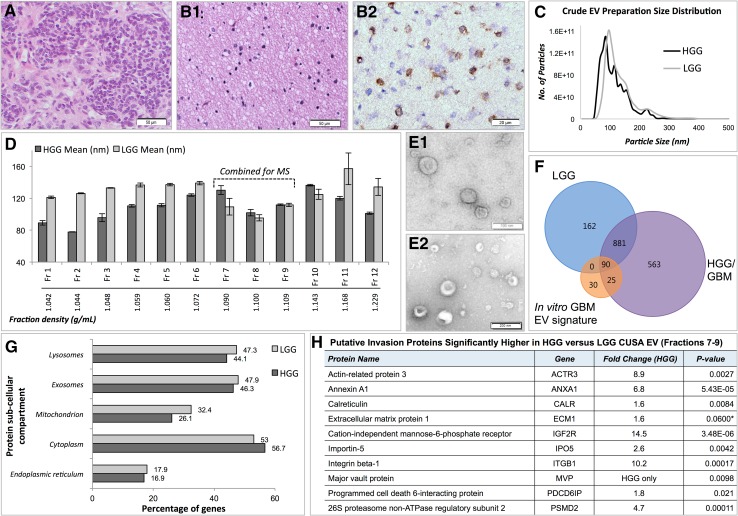
Extracellular vesicles (EVs) play key roles in glioblastoma (GBM) biology and represent novel sources of biomarkers that are detectable in the peripheral circulation. Despite this notionally non-invasive approach to assess GBM tumours in situ, a comprehensive GBM EV protein signature has not been described. Here, EVs secreted by six GBM cell lines were isolated and analysed by quantitative high-resolution mass spectrometry. Overall, 844 proteins were identified in the GBM EV proteome, of which 145 proteins were common to EVs secreted by all cell lines examined; included in the curated EV compendium (Vesiclepedia_559; http://microvesicles.org ). Levels of 14 EV proteins significantly correlated with cell invasion (invadopodia production; r2 > 0.5, p < 0.05), including several proteins that interact with molecules responsible for regulating invadopodia formation. Invadopodia, actin-rich membrane protrusions with proteolytic activity, are associated with more aggressive disease and are sites of EV release. Gene levels corresponding to invasion-related EV proteins showed that five genes (annexin A1, actin-related protein 3, integrin-β1, insulin-like growth factor 2 receptor and programmed cell death 6-interacting protein) were significantly higher in GBM tumours compared to normal brain in silico, with common functions relating to actin polymerisation and endosomal sorting. We also show that Cavitron Ultrasonic Surgical Aspirator (CUSA) washings are a novel source of brain tumour-derived EVs, demonstrated by particle tracking analysis, TEM and proteome profiling. Quantitative proteomics corroborated the high levels of proposed invasion-related proteins in EVs enriched from a GBM compared to low-grade astrocytoma tumour. Large-scale clinical follow-up of putative biomarkers, particularly the proposed survival marker annexin A1, is warranted.
Keywords: Annexin a1; Exosome; Extracellular vesicle; Glioblastoma; Invadopodia; Proteomics.
Figures
References
-
- Thery C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol. 2002;2:569–579. - PubMed
-
- Skog J, Wurdinger T, van Rijn S, Meijer DH, Gainche L, Sena-Esteves M, Curry WT, Jr., Carter BS, Krichevsky AM, Breakefield XO. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat Cell Biol. 2008;10(12):1470–1476. doi: 10.1038/ncb1800. - DOI - PMC - PubMed
-
- Kucharzewska P, Christianson HC, Welch JE, Svensson KJ, Fredlund E, Ringner M, Morgelin M, Bourseau-Guilmain E, Bengzon J, Belting M. Exosomes reflect the hypoxic status of glioma cells and mediate hypoxia-dependent activation of vascular cells during tumor development. PNAS. 2013;110(18):7312–7317. doi: 10.1073/pnas.1220998110. - DOI - PMC - PubMed
-
- Antonyak MA, Li B, Boroughs LK, Johnson JL, Druso JE, Bryant KL, Holowka DA, Cerione RA. Cancer cell-derived microvesicles induce transformation by transferring tissue transglutaminase and fibronectin to recipient cells. Proc Natl Acad Sci USA. 2011;108(12):4852–4857. doi: 10.1073/pnas.1017667108. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
