

Redox-dependent regulation of hepatocyte absent in melanoma 2 inflammasome activation in sterile liver injury in mice
- PMID: 27774630
- PMCID: PMC5191963
- DOI: 10.1002/hep.28893
Redox-dependent regulation of hepatocyte absent in melanoma 2 inflammasome activation in sterile liver injury in mice
Abstract
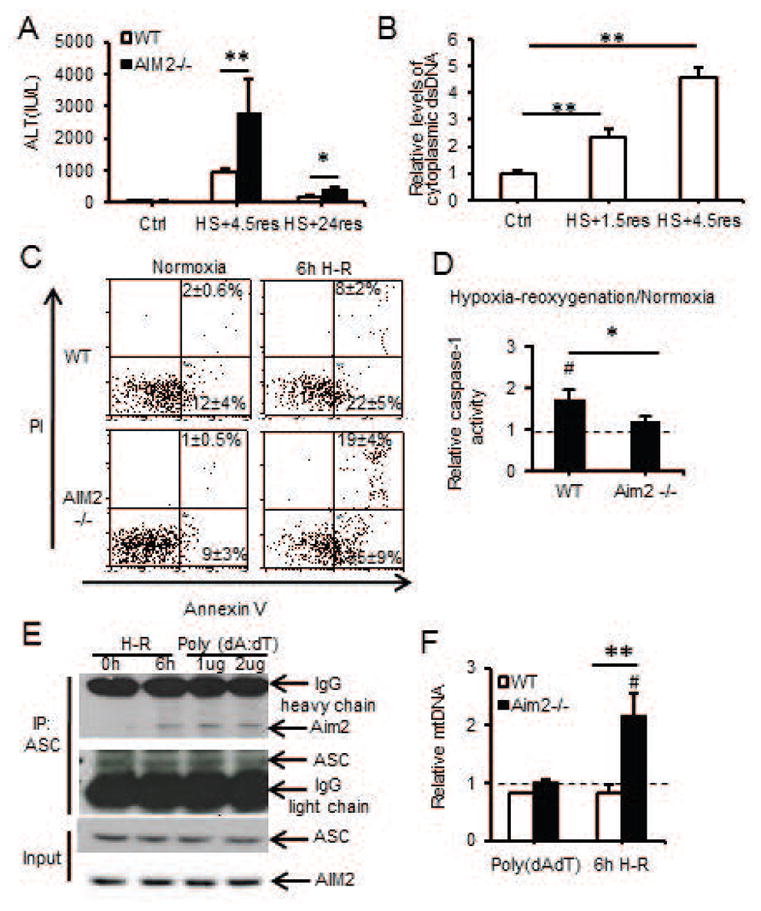
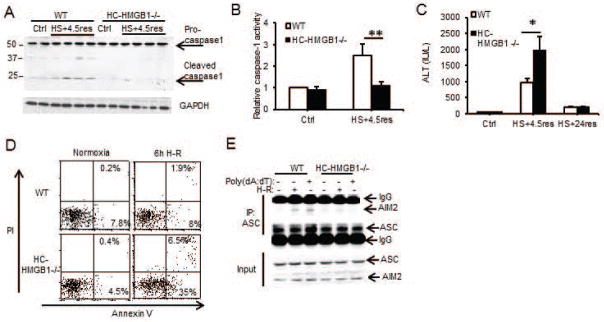
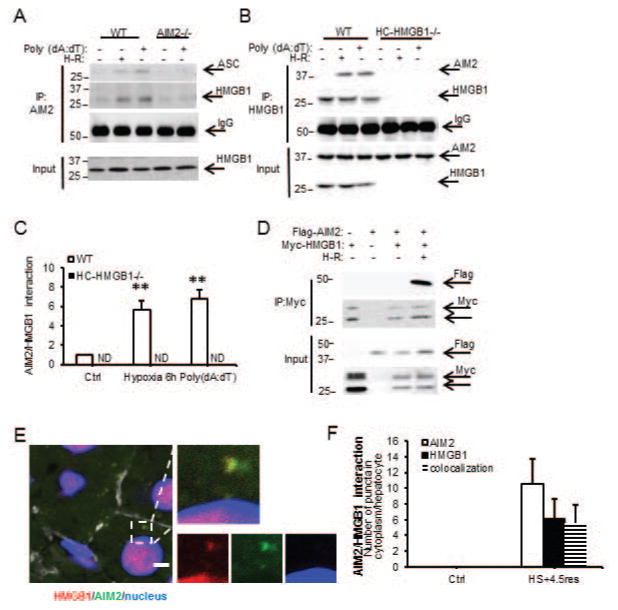
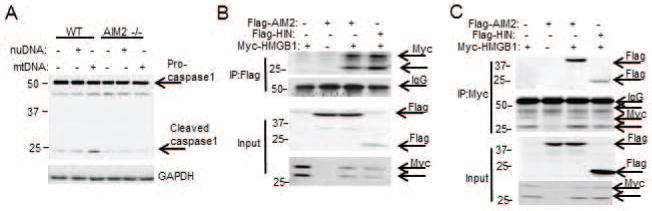
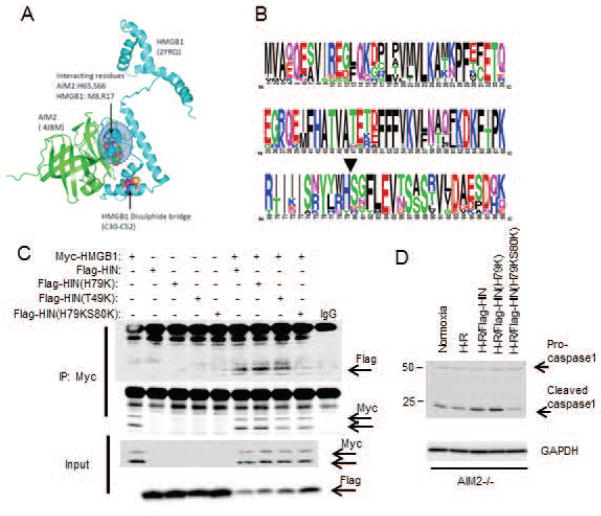
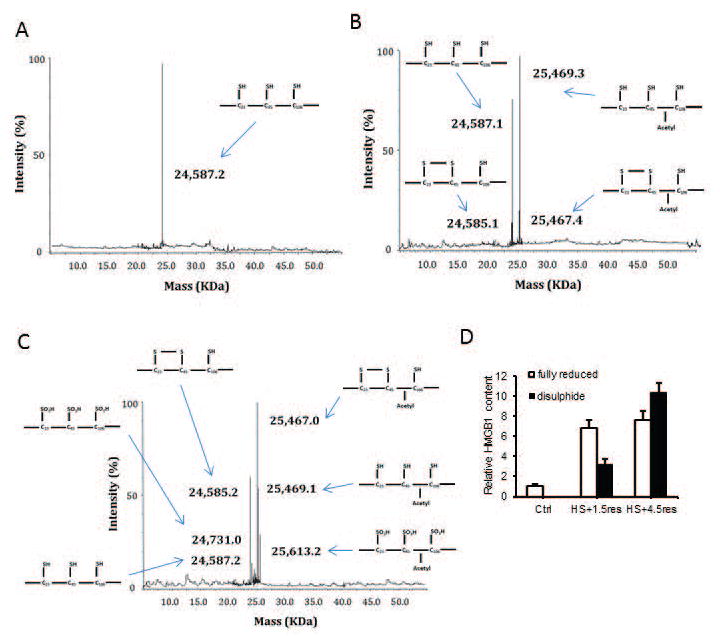
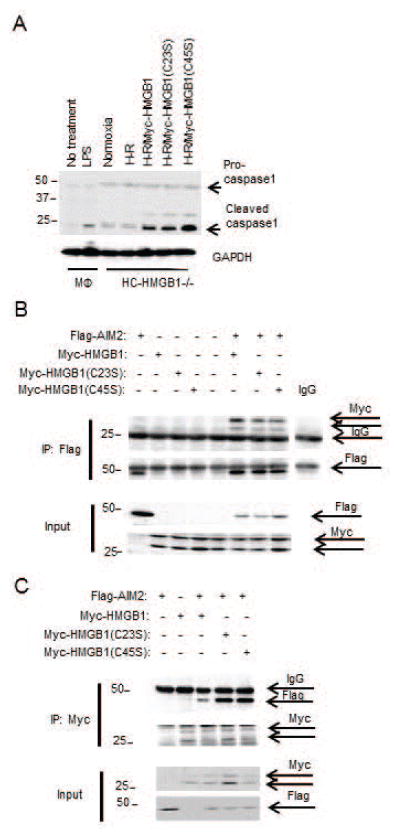
Sterile liver inflammation, such as liver ischemia-reperfusion, hemorrhagic shock after trauma, and drug-induced liver injury, is initiated and regulated by endogenous mediators including DNA and reactive oxygen species. Here, we identify a mechanism for redox-mediated regulation of absent in melanoma 2 (AIM2) inflammasome activation in hepatocytes after redox stress in mice, which occurs through interaction with cytosolic high mobility group box 1 (HMGB1). We show that in liver during hemorrhagic shock in mice and in hepatocytes after hypoxia with reoxygenation, cytosolic HMGB1 associates with AIM2 and is required for activation of caspase-1 in response to cytosolic DNA. Activation of caspase-1 through AIM2 leads to subsequent hepatoprotective responses such as autophagy. HMGB1 binds to AIM2 at a non-DNA-binding site on the hematopoietic interferon-inducible nuclear antigen domain of AIM2 to facilitate inflammasome and caspase-1 activation in hepatocytes. Furthermore, binding of HMGB1 to AIM2 is stronger with fully reduced all-thiol HMGB1 than with partially oxidized disulfide-HMGB1, and binding strength corresponds to caspase-1 activation. These data suggest that HMGB1 redox status regulates AIM2 inflammasome activation.
Conclusion: These findings suggest a novel and important mechanism for regulation of AIM2 inflammasome activation in hepatocytes during redox stress and may suggest broader implications for how this and other inflammasomes are activated and how their activation is regulated during cell stress, as well as the mechanisms of inflammasome regulation in nonimmune cell types. (Hepatology 2017;65:253-268).
© 2016 by the American Association for the Study of Liver Diseases.
Figures
Similar articles
-
Heatstroke induces liver injury via IL-1β and HMGB1-induced pyroptosis.J Hepatol. 2015 Sep;63(3):622-33. doi: 10.1016/j.jhep.2015.04.010. Epub 2015 Apr 27. J Hepatol. 2015. PMID: 25931416
-
Knockdown of the AIM2 molecule attenuates ischemia-reperfusion-induced spinal neuronal pyroptosis by inhibiting AIM2 inflammasome activation and subsequent release of cleaved caspase-1 and IL-1β.Neuropharmacology. 2019 Dec 1;160:107661. doi: 10.1016/j.neuropharm.2019.05.038. Epub 2019 Jun 8. Neuropharmacology. 2019. PMID: 31181224
-
Activation of NLRP3 and AIM2 inflammasomes in Kupffer cells in hepatic ischemia/reperfusion.FEBS J. 2015 Jan;282(2):259-70. doi: 10.1111/febs.13123. Epub 2014 Nov 14. FEBS J. 2015. PMID: 25327779
-
AIM2 inflammasome activation and regulation: A structural perspective.J Struct Biol. 2017 Dec;200(3):279-282. doi: 10.1016/j.jsb.2017.08.001. Epub 2017 Aug 13. J Struct Biol. 2017. PMID: 28813641 Free PMC article. Review.
-
The Emerging Relevance of AIM2 in Liver Disease.Int J Mol Sci. 2020 Sep 7;21(18):6535. doi: 10.3390/ijms21186535. Int J Mol Sci. 2020. PMID: 32906750 Free PMC article. Review.
Cited by
-
TSLP protects against liver I/R injury via activation of the PI3K/Akt pathway.JCI Insight. 2019 Nov 14;4(22):e129013. doi: 10.1172/jci.insight.129013. JCI Insight. 2019. PMID: 31723054 Free PMC article.
-
Inflammatory Caspase Activity Mediates HMGB1 Release and Differentiation in Myoblasts Affected by Peripheral Arterial Disease.Cells. 2022 Mar 30;11(7):1163. doi: 10.3390/cells11071163. Cells. 2022. PMID: 35406727 Free PMC article.
-
Traumatic Brain Injury-Induced Acute Lung Injury: Evidence for Activation and Inhibition of a Neural-Respiratory-Inflammasome Axis.J Neurotrauma. 2018 Sep 1;35(17):2067-2076. doi: 10.1089/neu.2017.5430. Epub 2018 Jun 8. J Neurotrauma. 2018. PMID: 29648974 Free PMC article.
-
"Thinking" vs. "Talking": Differential Autocrine Inflammatory Networks in Isolated Primary Hepatic Stellate Cells and Hepatocytes under Hypoxic Stress.Front Physiol. 2017 Dec 22;8:1104. doi: 10.3389/fphys.2017.01104. eCollection 2017. Front Physiol. 2017. PMID: 29312006 Free PMC article.
-
Inflammasome and autophagy regulation - a two-way street.Mol Med. 2017 Sep;23:188-195. doi: 10.2119/molmed.2017.00077. Epub 2017 Jul 24. Mol Med. 2017. PMID: 28741645 Free PMC article. Review.
References
-
- Marques PE, Oliveira AG, Pereira RV, David BA, Gomides LF, Saraiva AM, Pires DA, et al. Hepatic DNA deposition drives drug-induced liver injury and inflammation in mice. Hepatology. 2015;61:348–360. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
