

Antibiotic effects on gut microbiota and metabolism are host dependent
- PMID: 27775551
- PMCID: PMC5127688
- DOI: 10.1172/JCI86674
Antibiotic effects on gut microbiota and metabolism are host dependent
Abstract
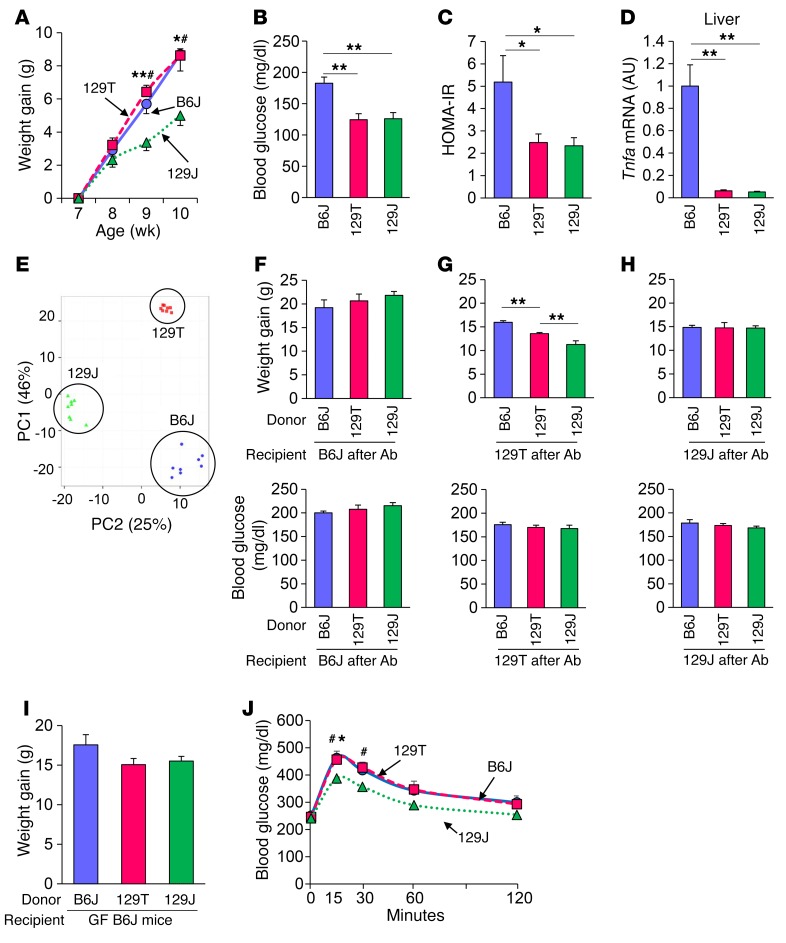
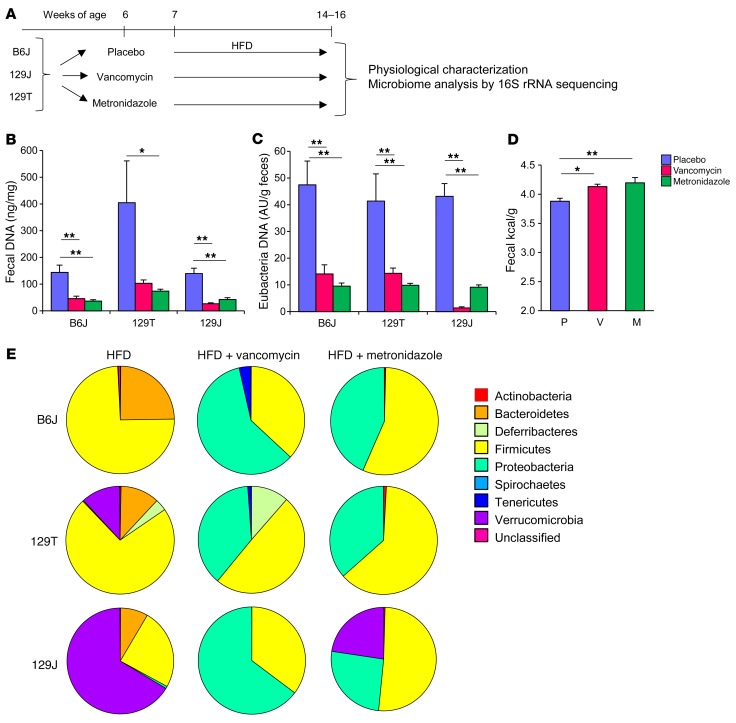
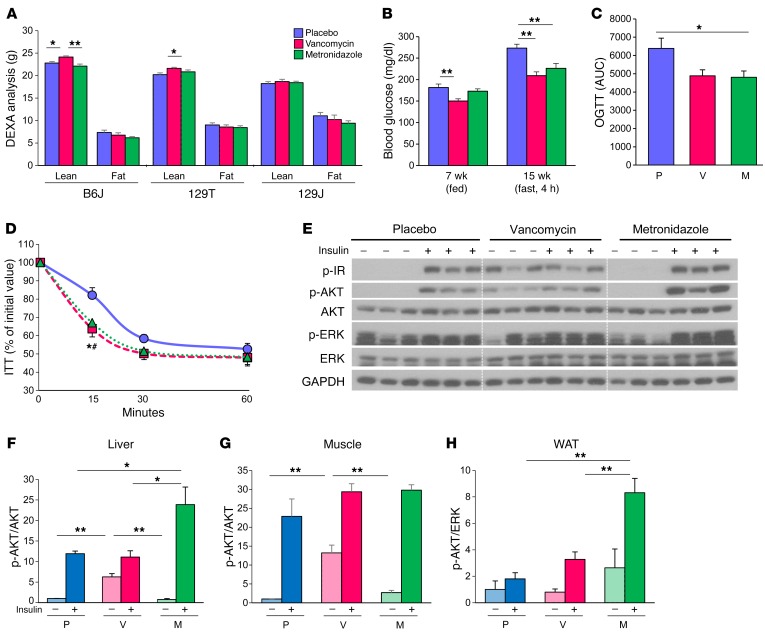
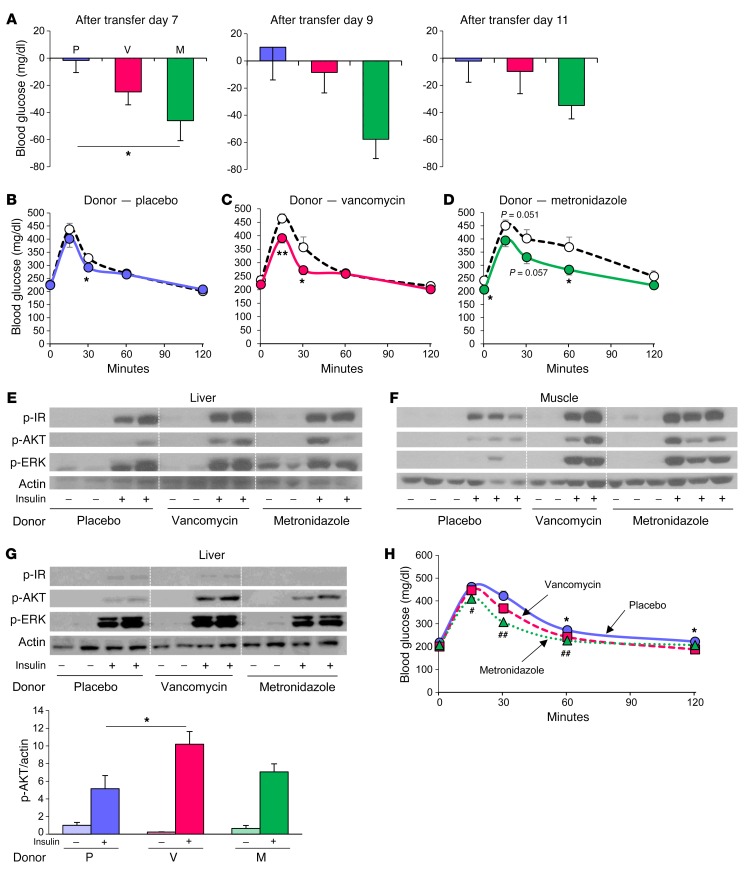
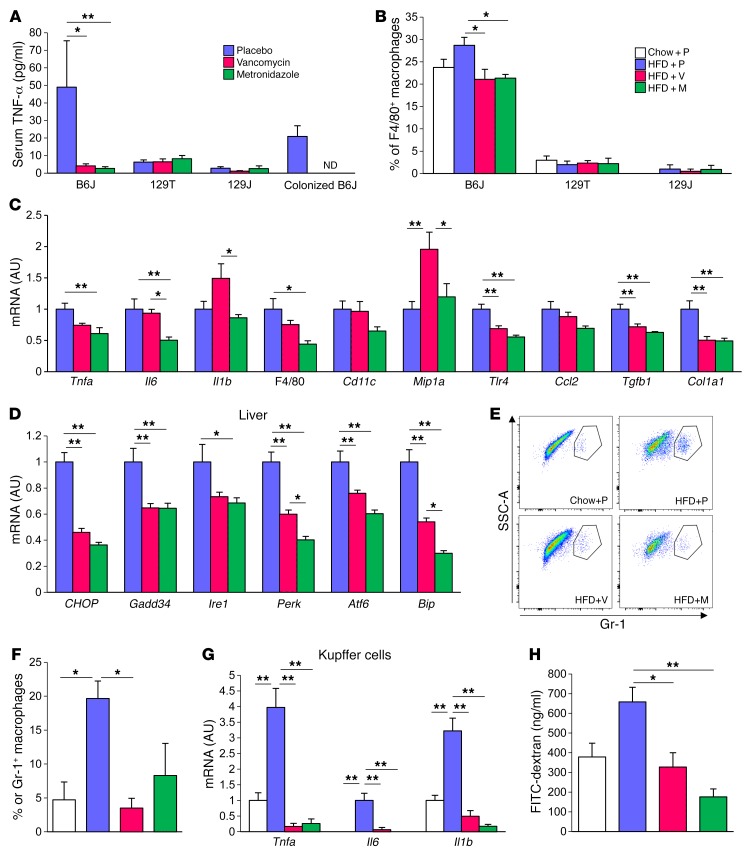
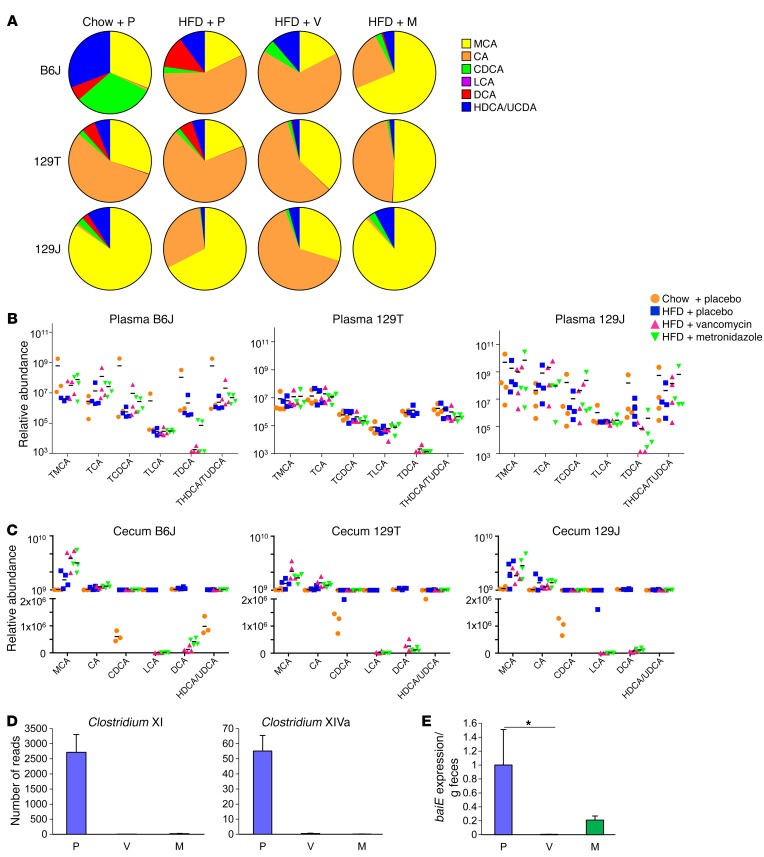
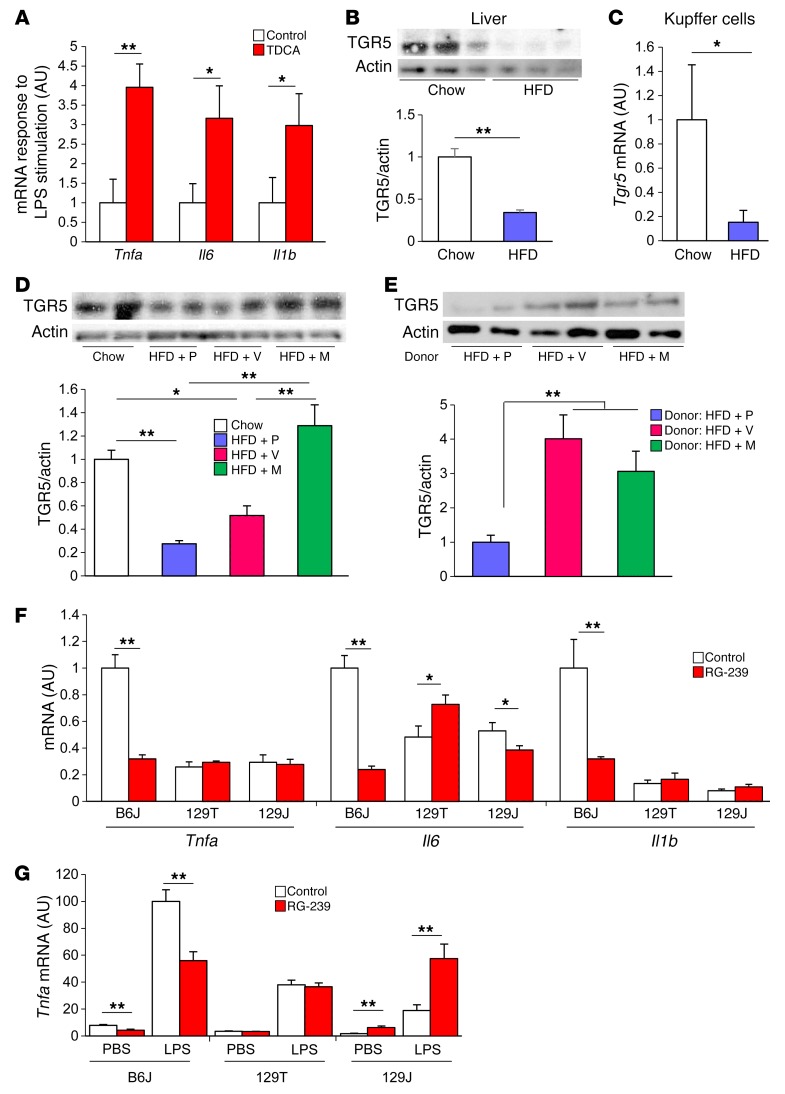
Interactions of diet, gut microbiota, and host genetics play important roles in the development of obesity and insulin resistance. Here, we have investigated the molecular links between gut microbiota, insulin resistance, and glucose metabolism in 3 inbred mouse strains with differing susceptibilities to metabolic syndrome using diet and antibiotic treatment. Antibiotic treatment altered intestinal microbiota, decreased tissue inflammation, improved insulin signaling in basal and stimulated states, and improved glucose metabolism in obesity- and diabetes-prone C57BL/6J mice on a high-fat diet (HFD). Many of these changes were reproduced by the transfer of gut microbiota from antibiotic-treated donors to germ-free or germ-depleted mice. These physiological changes closely correlated with changes in serum bile acids and levels of the antiinflammatory bile acid receptor Takeda G protein-coupled receptor 5 (TGR5) and were partially recapitulated by treatment with a TGR5 agonist. In contrast, antibiotic treatment of HFD-fed, obesity-resistant 129S1 and obesity-prone 129S6 mice did not improve metabolism, despite changes in microbiota and bile acids. These mice also failed to show a reduction in inflammatory gene expression in response to the TGR5 agonist. Thus, changes in bile acid and inflammatory signaling, insulin resistance, and glucose metabolism driven by an HFD can be modified by antibiotic-induced changes in gut microbiota; however, these effects depend on important interactions with the host's genetic background and inflammatory potential.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
