

Preadolescent Adversity Programs a Disrupted Maternal Stress Reactivity in Humans and Mice
- PMID: 27776734
- PMCID: PMC5326692
- DOI: 10.1016/j.biopsych.2016.08.027
Preadolescent Adversity Programs a Disrupted Maternal Stress Reactivity in Humans and Mice
Abstract
Background: Adverse childhood experiences (ACEs) are one of the greatest predictors of affective disorders for women. Periods of dynamic hormonal flux, including pregnancy, exacerbate the risk for affective disturbance and promote hypothalamic-pituitary-adrenal (HPA) axis dysregulation, a key feature of affective disorders. Little is understood as to how stress experienced in late childhood, defined as preadolescence, alters the programming unique to this period of brain maturation and its interaction with the hormonal changes of pregnancy and postpartum.
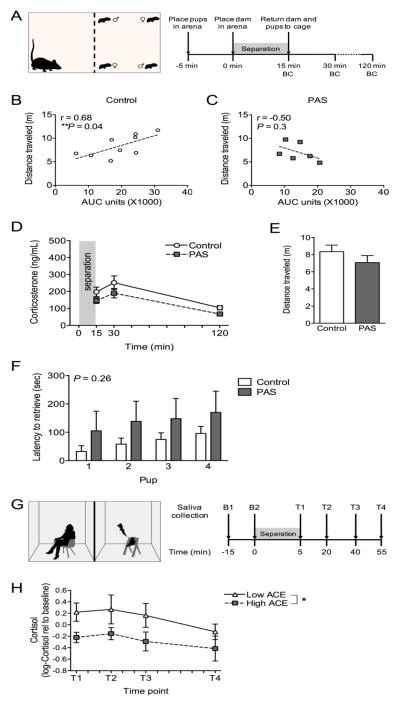
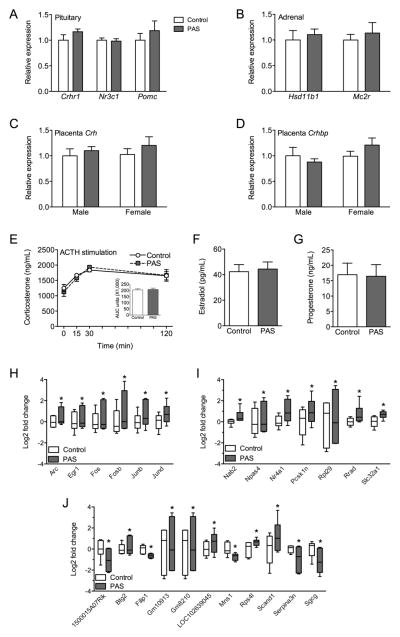
Methods: Preadolescent female mice were exposed to chronic stress and examined for changes in their HPA axis during pregnancy and postpartum, including assessment of maternal-specific stress responsiveness and transcriptomics of the paraventricular nucleus of the hypothalamus. Translationally, pregnant women with low or high ACEs were examined for their maternal stress responsiveness.
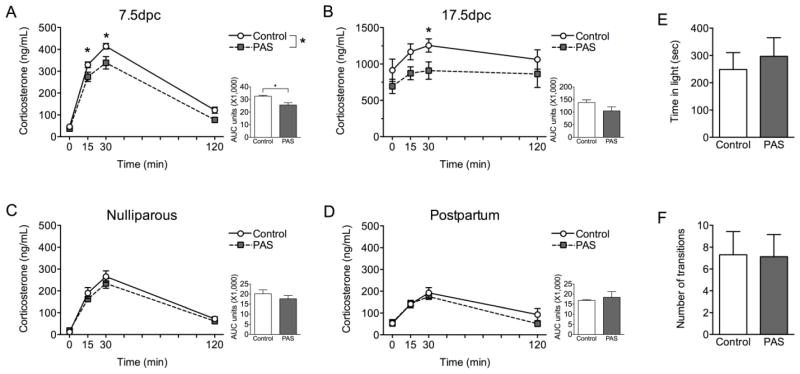
Results: As predicted, preadolescent stress in mice resulted in a significant blunting of the corticosterone response during pregnancy. Transcriptomic analysis of the paraventricular nucleus revealed widespread changes in expression of immediate early genes and their targets, supporting the likely involvement of an upstream epigenetic mechanism. Critically, in our human studies, the high ACE women showed a significant blunting of the HPA response.
Conclusions: This unique mouse model recapitulates a clinical outcome of a hyporesponsive HPA stress axis, an important feature of affective disorders, during a dynamic hormonal period, and suggests involvement of transcriptional regulation in the hypothalamus. These studies identify a novel mouse model of female ACEs that can be used to examine how additional life adversity may provoke disease risk or resilience.
Keywords: Adolescence; HPA axis; Paraventricular nucleus; Postpartum; Pregnancy; Stress.
Copyright © 2016 Society of Biological Psychiatry. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
The Nature of Nurture: How Developmental Experiences Program Adult Stress Circuitry.Biol Psychiatry. 2017 Apr 15;81(8):e57-e59. doi: 10.1016/j.biopsych.2017.02.004. Biol Psychiatry. 2017. PMID: 28317550 Free PMC article. No abstract available.
References
-
- Heim C, Shugart M, Craighead WE, Nemeroff CB. Neurobiological and psychiatric consequences of child abuse and neglect. Dev Psychobiol. 2010;52:671–90. - PubMed
-
- Janssen I, Krabbendam L, Bak M, Hanssen M, Vollebergh W, de Graaf R, van Os J. Childhood abuse as a risk factor for psychotic experiences. Acta Psychiatr Scand. 2004;109:38–45. - PubMed
-
- Chapman DP, Whitfield CL, Felitti VJ, Dube SR, Edwards VJ, Anda RF. Adverse childhood experiences and the risk of depressive disorders in adulthood. J Affect Disord. 2004;82:217–25. - PubMed
-
- McCauley J, Kern DE, Kolodner K, Dill L, Schroeder AF, DeChant HK, et al. Clinical characteristics of women with a history of childhood abuse: unhealed wounds. JAMA. 1997;277:1362–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AG048839/AG/NIA NIH HHS/United States
- K23 MH107831/MH/NIMH NIH HHS/United States
- R01 MH087597/MH/NIMH NIH HHS/United States
- K23 MH092399/MH/NIMH NIH HHS/United States
- K24 DA030301/DA/NIDA NIH HHS/United States
- K12 HD085848/HD/NICHD NIH HHS/United States
- R01 MH073030/MH/NIMH NIH HHS/United States
- R21 MH104184/MH/NIMH NIH HHS/United States
- R33 MH104184/MH/NIMH NIH HHS/United States
- P50 MH099910/MH/NIMH NIH HHS/United States
- R37 MH108286/MH/NIMH NIH HHS/United States
- R01 MH091258/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
