

Snap Your Fingers! An ERP/sLORETA Study Investigating Implicit Processing of Self- vs. Other-Related Movement Sounds Using the Passive Oddball Paradigm
- PMID: 27777557
- PMCID: PMC5056175
- DOI: 10.3389/fnhum.2016.00465
Snap Your Fingers! An ERP/sLORETA Study Investigating Implicit Processing of Self- vs. Other-Related Movement Sounds Using the Passive Oddball Paradigm
Abstract
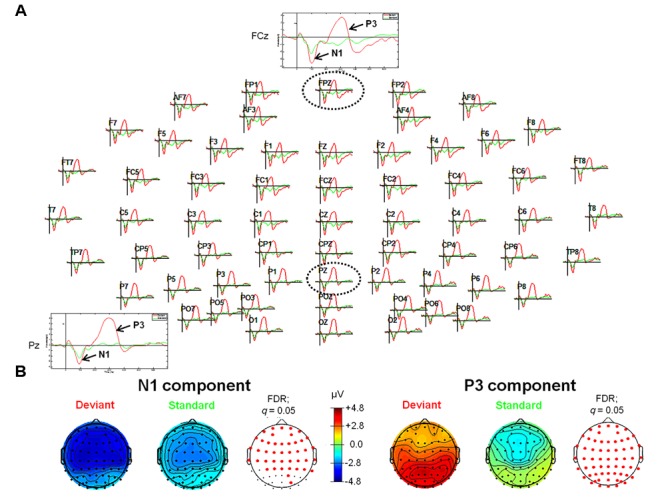
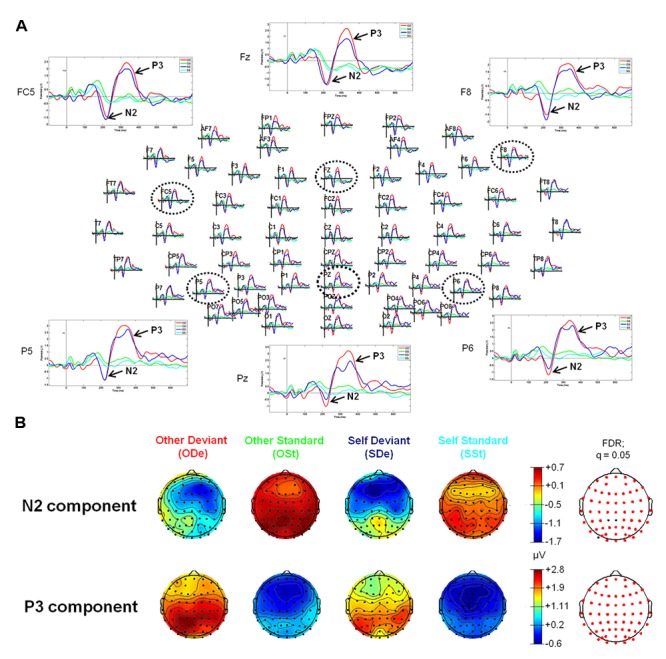
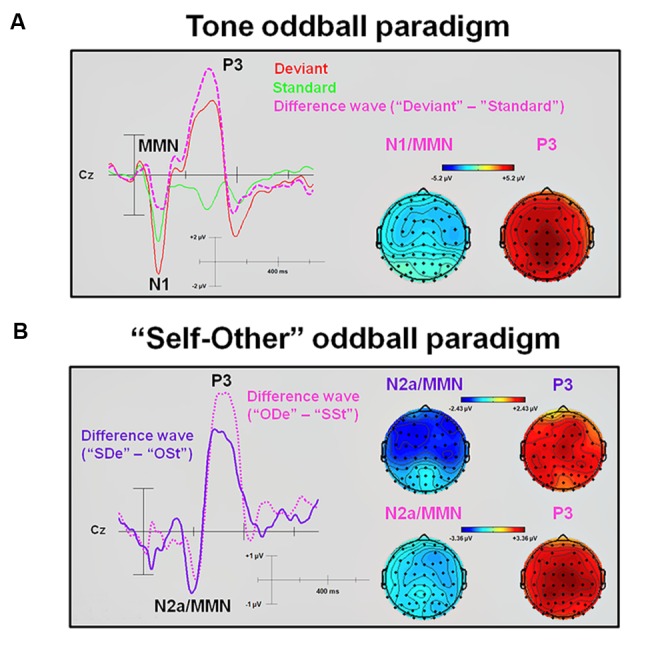
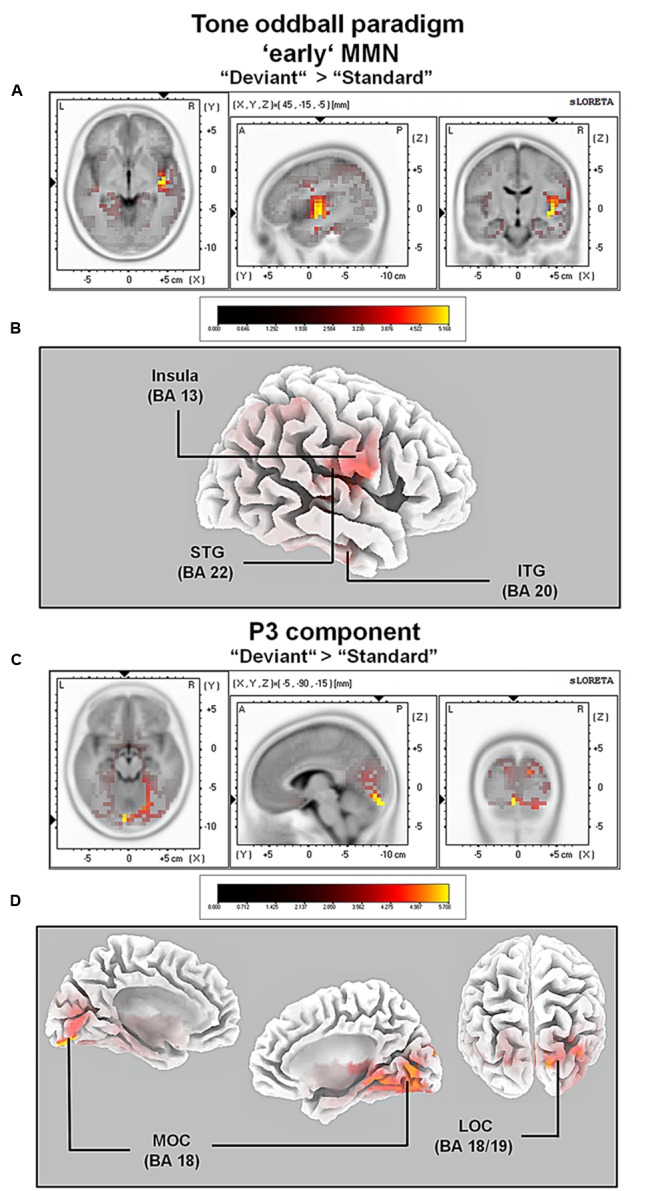
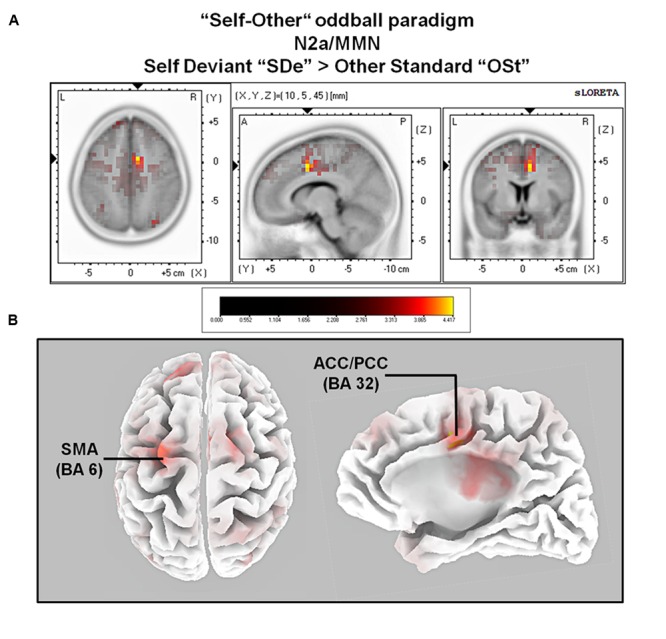
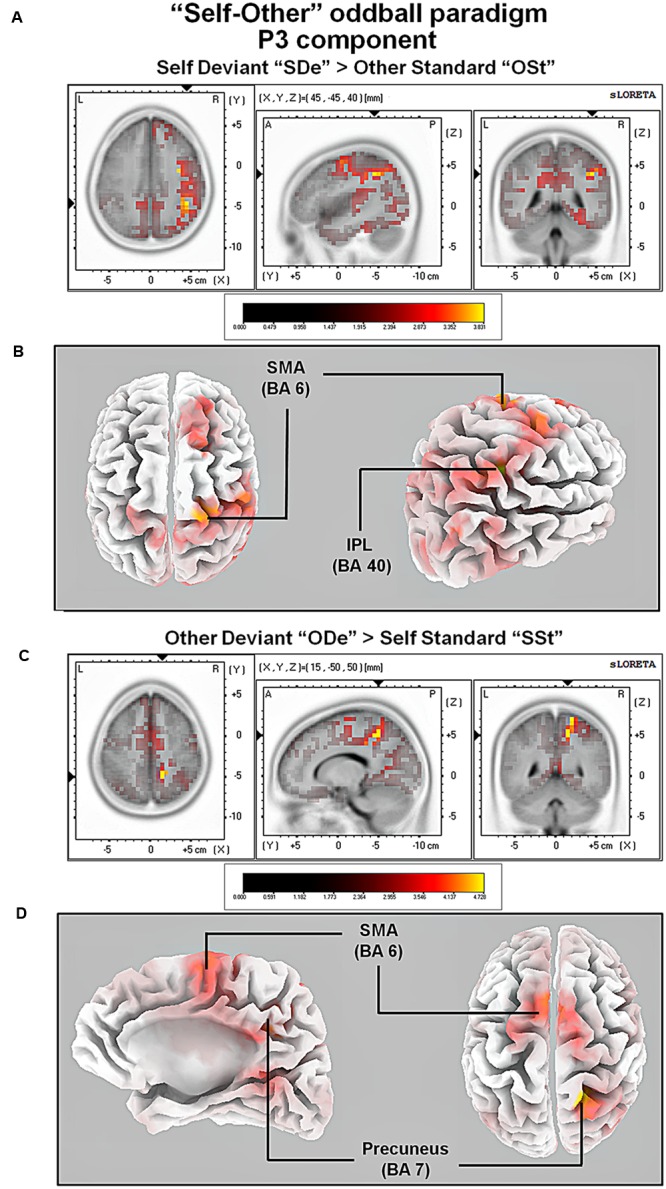
So far, neurophysiological studies have investigated implicit and explicit self-related processing particularly for self-related stimuli such as the own face or name. The present study extends previous research to the implicit processing of self-related movement sounds and explores their spatio-temporal dynamics. Event-related potentials (ERPs) were assessed while participants (N = 12 healthy subjects) listened passively to previously recorded self- and other-related finger snapping sounds, presented either as deviants or standards during an oddball paradigm. Passive listening to low (500 Hz) and high (1000 Hz) pure tones served as additional control. For self- vs. other-related finger snapping sounds, analysis of ERPs revealed significant differences in the time windows of the N2a/MMN and P3. An subsequent source localization analysis with standardized low-resolution brain electromagnetic tomography (sLORETA) revealed increased cortical activation in distinct motor areas such as the supplementary motor area (SMA) in the N2a/mismatch negativity (MMN) as well as the P3 time window during processing of self- and other-related finger snapping sounds. In contrast, brain regions associated with self-related processing [e.g., right anterior/posterior cingulate cortex (ACC/PPC)] as well as the right inferior parietal lobule (IPL) showed increased activation particularly during processing of self- vs. other-related finger snapping sounds in the time windows of the N2a/MMN (ACC/PCC) or the P3 (IPL). None of these brain regions showed enhanced activation while listening passively to low (500 Hz) and high (1000 Hz) pure tones. Taken together, the current results indicate (1) a specific role of motor regions such as SMA during auditory processing of movement-related information, regardless of whether this information is self- or other-related, (2) activation of neural sources such as the ACC/PCC and the IPL during implicit processing of self-related movement stimuli, and (3) their differential temporal activation during deviance (N2a/MMN - ACC/PCC) and target detection (P3 - IPL) of self- vs. other-related movement sounds.
Keywords: EEG; N2a/MMN; P3; finger snapping sounds; source localization.
Figures
Similar articles
-
The spatio-temporal dynamics of deviance and target detection in the passive and active auditory oddball paradigm: a sLORETA study.BMC Neurosci. 2018 Apr 19;19(1):25. doi: 10.1186/s12868-018-0422-3. BMC Neurosci. 2018. PMID: 29673322 Free PMC article.
-
Self vs. other: neural correlates underlying agent identification based on unimodal auditory information as revealed by electrotomography (sLORETA).Neuroscience. 2014 Feb 14;259:25-34. doi: 10.1016/j.neuroscience.2013.11.042. Epub 2013 Dec 1. Neuroscience. 2014. PMID: 24295635
-
Implicit auditory perception of local and global irregularities in passive listening condition.Neuropsychologia. 2022 Jan 28;165:108129. doi: 10.1016/j.neuropsychologia.2021.108129. Epub 2021 Dec 18. Neuropsychologia. 2022. PMID: 34929262
-
Neural substrates of normal and impaired preattentive sensory discrimination in large cohorts of nonpsychiatric subjects and schizophrenia patients as indexed by MMN and P3a change detection responses.Neuroimage. 2013 Feb 1;66:594-603. doi: 10.1016/j.neuroimage.2012.09.074. Epub 2012 Oct 22. Neuroimage. 2013. PMID: 23085112 Free PMC article.
-
Stimulus-dependent activations and attention-related modulations in the auditory cortex: a meta-analysis of fMRI studies.Hear Res. 2014 Jan;307:29-41. doi: 10.1016/j.heares.2013.08.001. Epub 2013 Aug 11. Hear Res. 2014. PMID: 23938208 Review.
Cited by
-
The spatio-temporal dynamics of deviance and target detection in the passive and active auditory oddball paradigm: a sLORETA study.BMC Neurosci. 2018 Apr 19;19(1):25. doi: 10.1186/s12868-018-0422-3. BMC Neurosci. 2018. PMID: 29673322 Free PMC article.
-
The ultrafast snap of a finger is mediated by skin friction.J R Soc Interface. 2021 Nov;18(184):20210672. doi: 10.1098/rsif.2021.0672. Epub 2021 Nov 17. J R Soc Interface. 2021. PMID: 34784775 Free PMC article.
-
Neurophysiological Oscillatory Mechanisms Underlying the Effect of Mirror Visual Feedback-Induced Illusion of Hand Movements on Nociception and Cortical Activation.Brain Sci. 2024 Jul 12;14(7):696. doi: 10.3390/brainsci14070696. Brain Sci. 2024. PMID: 39061436 Free PMC article.
-
A Signature of Passivity? An Explorative Study of the N3 Event- Related Potential Component in Passive Oddball Tasks.Front Neurosci. 2019 Apr 24;13:365. doi: 10.3389/fnins.2019.00365. eCollection 2019. Front Neurosci. 2019. PMID: 31068776 Free PMC article.
-
Processing Self-Related Information Under Non-attentional Conditions Revealed by Visual MMN.Front Hum Neurosci. 2022 Apr 6;16:782496. doi: 10.3389/fnhum.2022.782496. eCollection 2022. Front Hum Neurosci. 2022. PMID: 35463934 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources