

The actin binding proteins cortactin and HS1 are dispensable for platelet actin nodule and megakaryocyte podosome formation
- PMID: 27778524
- PMCID: PMC5274539
- DOI: 10.1080/09537104.2016.1235688
The actin binding proteins cortactin and HS1 are dispensable for platelet actin nodule and megakaryocyte podosome formation
Abstract
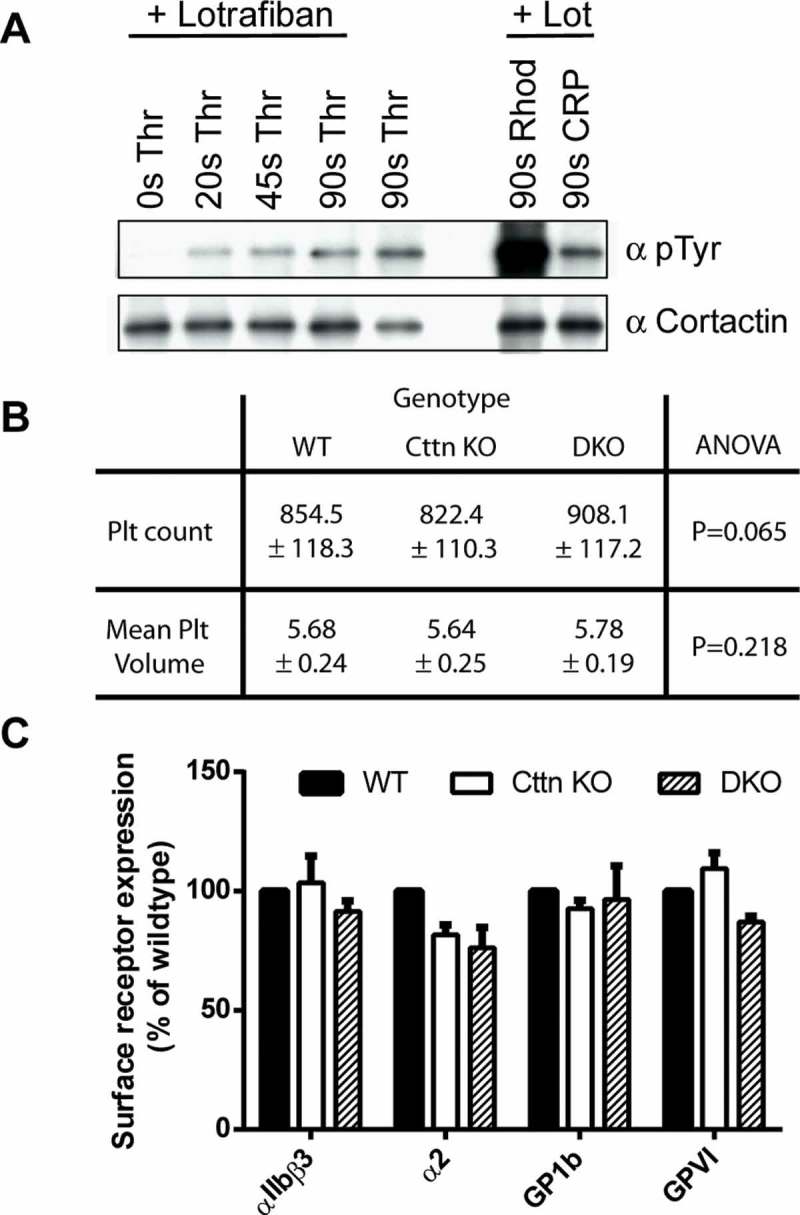
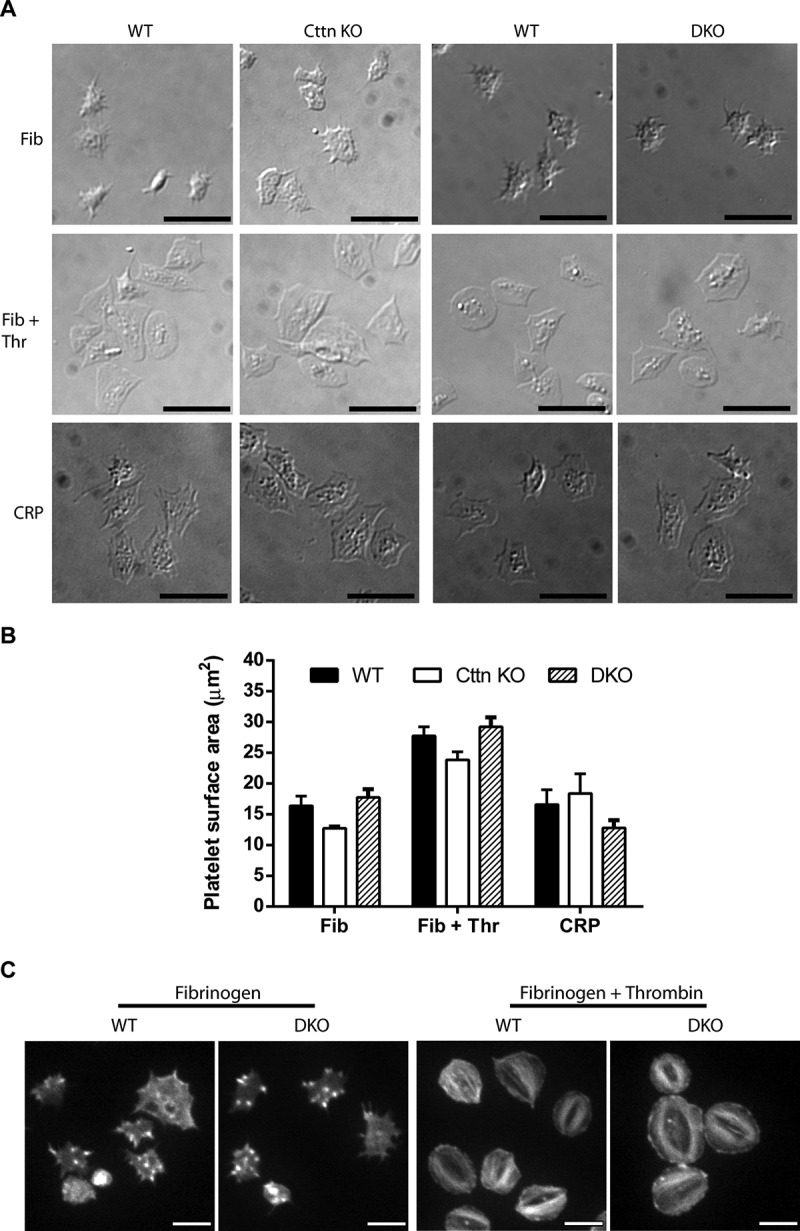
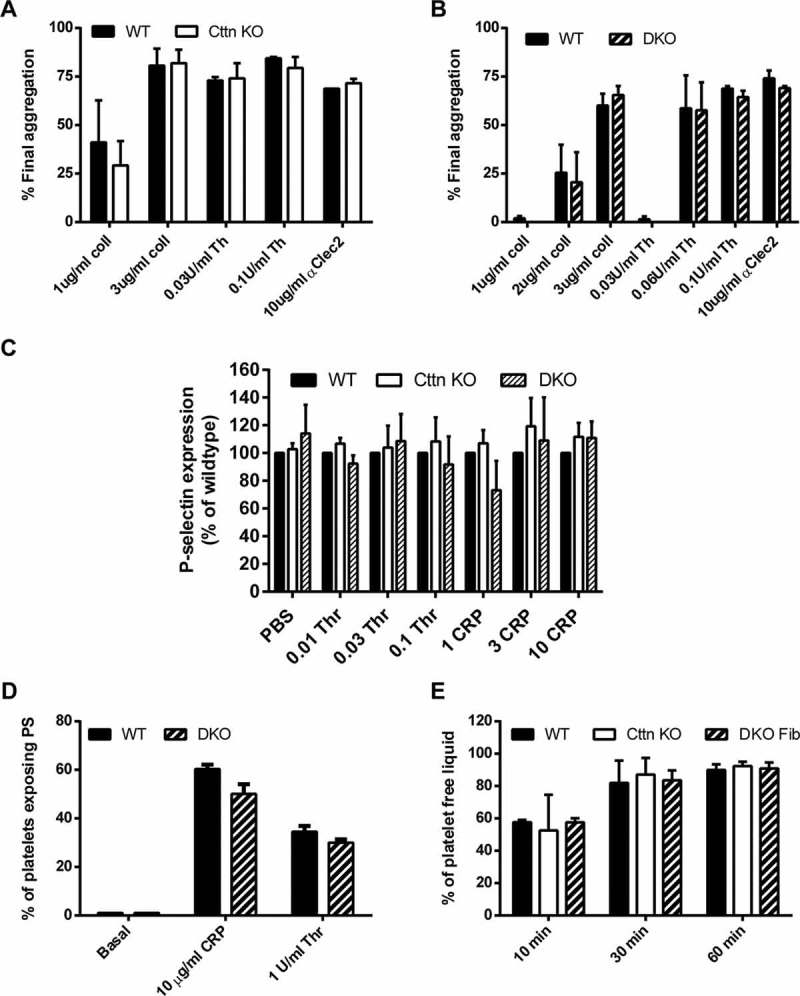
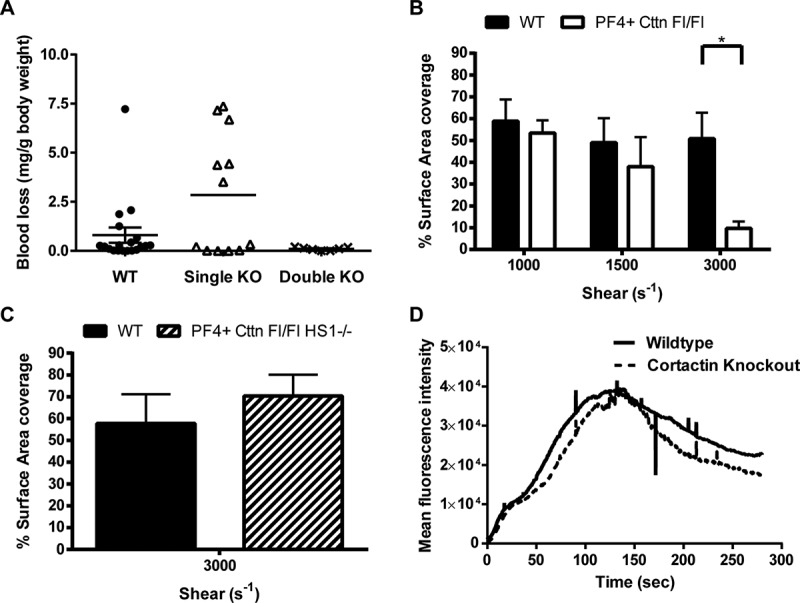
A dynamic, properly organised actin cytoskeleton is critical for the production and haemostatic function of platelets. The Wiskott Aldrich Syndrome protein (WASp) and Actin-Related Proteins 2 & 3 Complex (Arp2/3 complex) are critical mediators of actin polymerisation and organisation in many cell types. In platelets and megakaryocytes, these proteins have been shown to be important for proper platelet production and function. The cortactin family of proteins (Cttn & HS1) are known to regulate WASp-Arp2/3-mediated actin polymerisation in other cell types and so here we address the role of these proteins in platelets using knockout mouse models. We generated mice lacking Cttn and HS1 in the megakaryocyte/platelet lineage. These mice had normal platelet production, with platelet number, size and surface receptor profile comparable to controls. Platelet function was also unaffected by loss of Cttn/HS1 with no differences observed in a range of platelet function assays including aggregation, secretion, spreading, clot retraction or tyrosine phosphorylation. No effect on tail bleeding time or in thrombosis models was observed. In addition, platelet actin nodules, and megakaryocyte podosomes, actin-based structures known to be dependent on WASp and the Arp2/3 complex, formed normally. We conclude that despite the importance of WASp and the Arp2/3 complex in regulating F-actin dynamics in many cells types, the role of cortactin in their regulation appears to be fulfilled by other proteins in platelets.
Keywords: Actin nodules; Cortactin; HS1; platelets; podosomes.
Figures
Similar articles
-
Hematopoietic lineage cell-specific protein 1 functions in concert with the Wiskott-Aldrich syndrome protein to promote podosome array organization and chemotaxis in dendritic cells.J Immunol. 2011 Apr 15;186(8):4805-18. doi: 10.4049/jimmunol.1003102. Epub 2011 Mar 11. J Immunol. 2011. PMID: 21398607 Free PMC article.
-
Platelet actin nodules are podosome-like structures dependent on Wiskott-Aldrich syndrome protein and ARP2/3 complex.Nat Commun. 2015 Jun 1;6:7254. doi: 10.1038/ncomms8254. Nat Commun. 2015. PMID: 26028144 Free PMC article.
-
Studies on the actin-binding protein HS1 in platelets.BMC Cell Biol. 2007 Nov 9;8:46. doi: 10.1186/1471-2121-8-46. BMC Cell Biol. 2007. PMID: 17996076 Free PMC article.
-
Formin proteins in megakaryocytes and platelets: regulation of actin and microtubule dynamics.Platelets. 2019;30(1):23-30. doi: 10.1080/09537104.2018.1481937. Epub 2018 Jun 18. Platelets. 2019. PMID: 29913076 Free PMC article. Review.
-
Cortactin function in invadopodia.Small GTPases. 2020 Jul;11(4):256-270. doi: 10.1080/21541248.2017.1405773. Epub 2017 Dec 31. Small GTPases. 2020. PMID: 29172953 Free PMC article. Review.
Cited by
-
Hematopoietic lineage cell-specific protein 1 (HS1), a hidden player in migration, invasion, and tumor formation, is over-expressed in ovarian carcinoma cells.Oncotarget. 2018 Aug 24;9(66):32609-32623. doi: 10.18632/oncotarget.25975. eCollection 2018 Aug 24. Oncotarget. 2018. PMID: 30220969 Free PMC article.
-
Critical role of the HDAC6-cortactin axis in human megakaryocyte maturation leading to a proplatelet-formation defect.Nat Commun. 2017 Nov 27;8(1):1786. doi: 10.1038/s41467-017-01690-2. Nat Commun. 2017. PMID: 29176689 Free PMC article.
-
Cortactin stabilization of actin requires actin-binding repeats and linker, is disrupted by specific substitutions, and is independent of nucleotide state.J Biol Chem. 2018 Aug 24;293(34):13022-13032. doi: 10.1074/jbc.RA118.004068. Epub 2018 Jun 21. J Biol Chem. 2018. PMID: 29929984 Free PMC article.
-
Alterations in Platelet Alpha-Granule Secretion and Adhesion on Collagen under Flow in Mice Lacking the Atypical Rho GTPase RhoBTB3.Cells. 2019 Feb 11;8(2):149. doi: 10.3390/cells8020149. Cells. 2019. PMID: 30754723 Free PMC article.
-
Role of Rho-GTPases in megakaryopoiesis.Small GTPases. 2021 Sep-Nov;12(5-6):399-415. doi: 10.1080/21541248.2021.1885134. Epub 2021 Feb 11. Small GTPases. 2021. PMID: 33570449 Free PMC article. Review.
References
-
- Poulter NS, Thomas SG.. Cytoskeletal regulation of platelet formation: Coordination of F-actin and microtubules. Int J Biochem Cell Biol 2015;66:69–74. - PubMed
-
- Sorrentino S, Studt JD, Medalia O, Tanuj Sapra K. Roll, adhere, spread and contract: structural mechanics of platelet function. Eur J Cell Biol 2015;94(3–4):129–138. - PubMed
-
- Bender M, Eckly A, Hartwig JH, Elvers M, Pleines I, Gupta S, Krohne G, Jeanclos E, Gohla A, Gurniak C, et al. ADF/n-cofilin-dependent actin turnover determines platelet formation and sizing. Blood 2010;116(10):1767–1775. - PubMed
-
- Bender M, Stritt S, Nurden P, van Eeuwijk JMM, Zieger B, Kentouche K, Schulze H, Morbach H, Stegner D, Heinze KG, et al. Megakaryocyte-specific Profilin1-deficiency alters microtubule stability and causes a Wiskott–Aldrich syndrome-like platelet defect. Nat Commun 2014;5:4746. - PubMed
-
- Léon C, Eckly A, Hechler B, Aleil B, Freund M, Ravanat C, Jourdain M, Nonne C, Weber J, Tiedt R, et al. Megakaryocyte-restricted MYH9 inactivation dramatically affects hemostasis while preserving platelet aggregation and secretion. Blood 2007;110(9):3183–3191. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous