

Differential roles of two delayed rectifier potassium currents in regulation of ventricular action potential duration and arrhythmia susceptibility
- PMID: 27779762
- PMCID: PMC5374112
- DOI: 10.1113/JP273191
Differential roles of two delayed rectifier potassium currents in regulation of ventricular action potential duration and arrhythmia susceptibility
Abstract
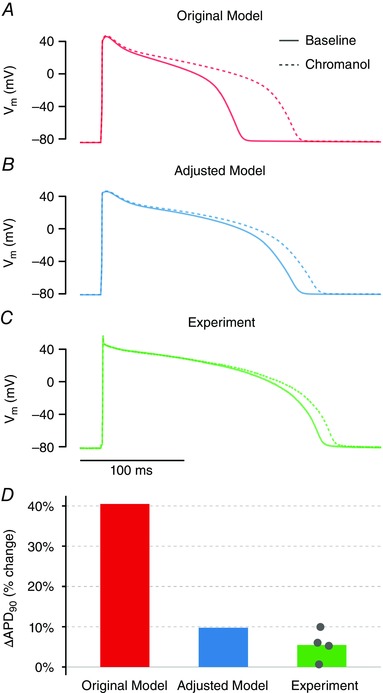
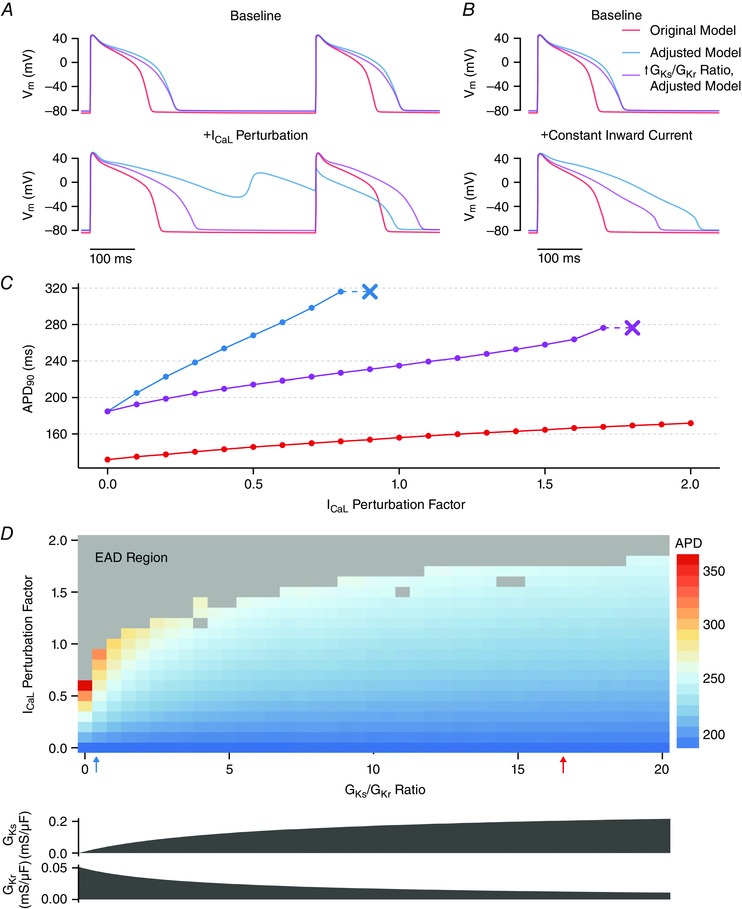
Key points: Arrhythmias result from disruptions to cardiac electrical activity, although the factors that control cellular action potentials are incompletely understood. We combined mathematical modelling with experiments in heart cells from guinea pigs to determine how cellular electrical activity is regulated. A mismatch between modelling predictions and the experimental results allowed us to construct an improved, more predictive mathematical model. The balance between two particular potassium currents dictates how heart cells respond to perturbations and their susceptibility to arrhythmias.
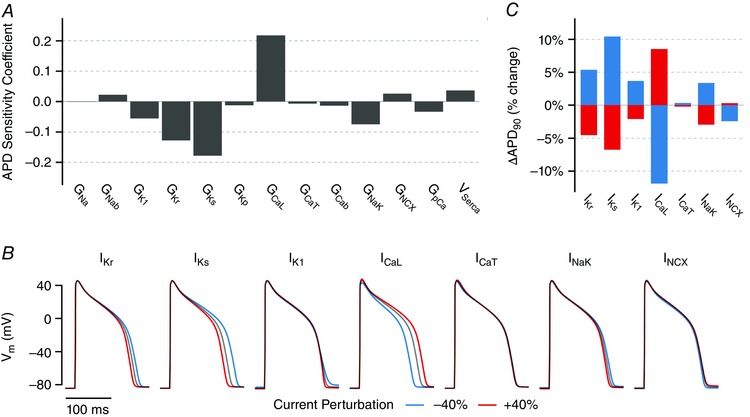
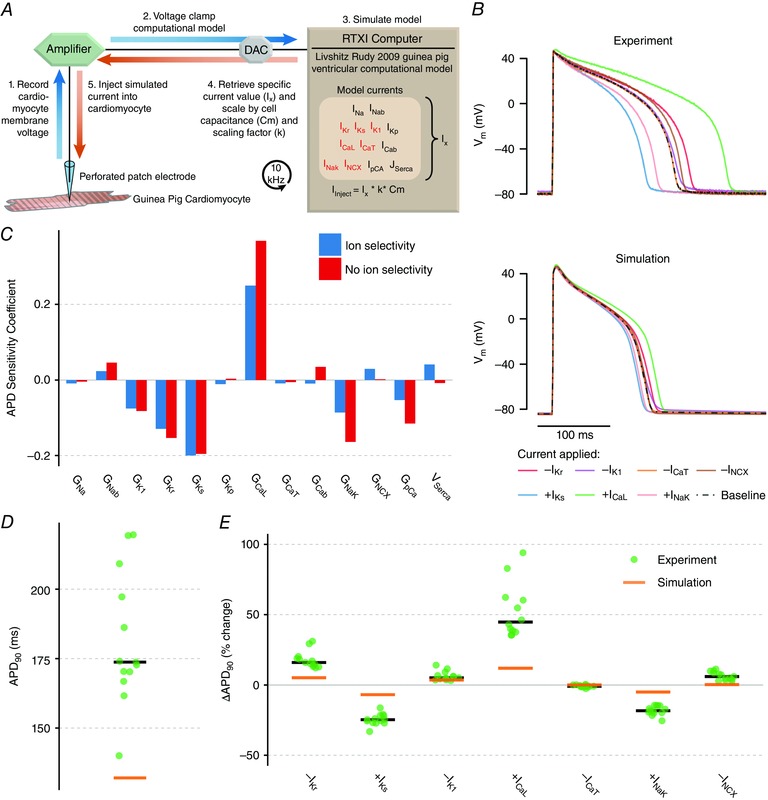
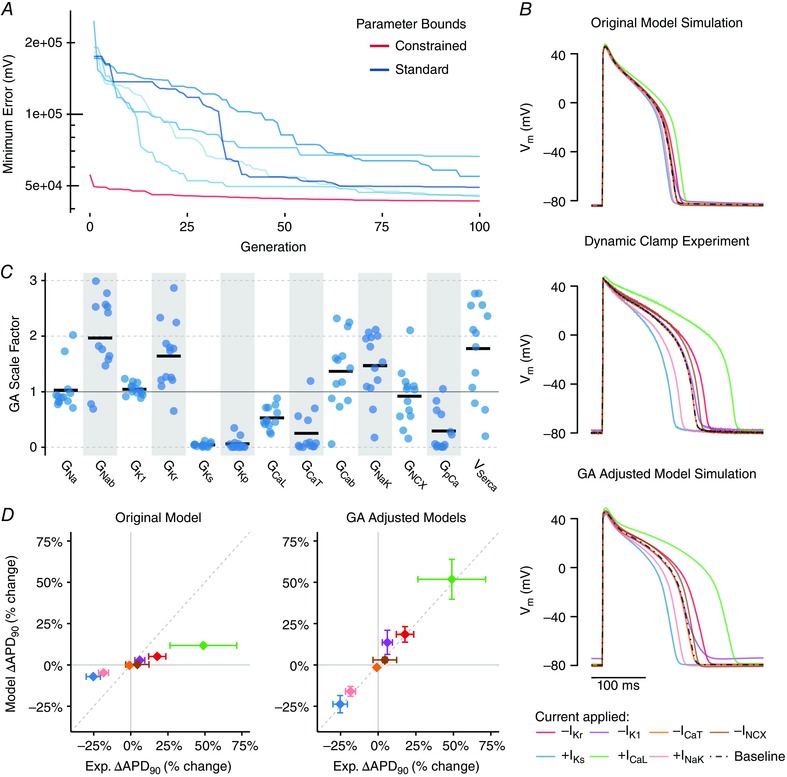
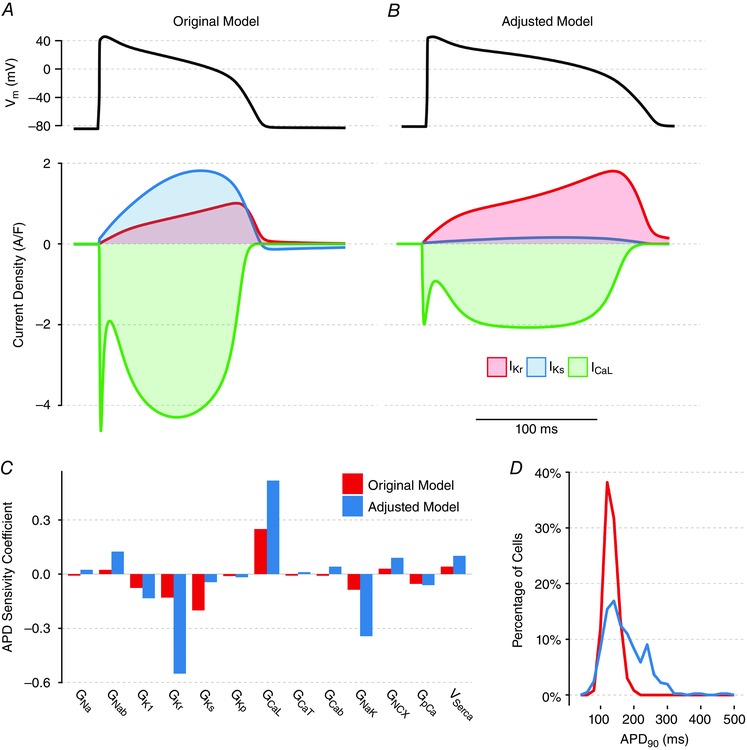
Abstract: Imbalances of ionic currents can destabilize the cardiac action potential and potentially trigger lethal cardiac arrhythmias. In the present study, we combined mathematical modelling with information-rich dynamic clamp experiments to determine the regulation of action potential morphology in guinea pig ventricular myocytes. Parameter sensitivity analysis was used to predict how changes in ionic currents alter action potential duration, and these were tested experimentally using dynamic clamp, a technique that allows for multiple perturbations to be tested in each cell. Surprisingly, we found that a leading mathematical model, developed with traditional approaches, systematically underestimated experimental responses to dynamic clamp perturbations. We then re-parameterized the model using a genetic algorithm, which allowed us to estimate ionic current levels in each of the cells studied. This unbiased model adjustment consistently predicted an increase in the rapid delayed rectifier K+ current and a drastic decrease in the slow delayed rectifier K+ current, and this prediction was validated experimentally. Subsequent simulations with the adjusted model generated the clinically relevant prediction that the slow delayed rectifier is better able to stabilize the action potential and suppress pro-arrhythmic events than the rapid delayed rectifier. In summary, iterative coupling of simulations and experiments enabled novel insight into how the balance between cardiac K+ currents influences ventricular arrhythmia susceptibility.
Keywords: ion channels; mathematical model; patch clamp.
© 2016 The Authors. The Journal of Physiology © 2016 The Physiological Society.
Figures
References
-
- Banyasz T, Horvath B, Virag L, Barandi L, Szentandrassy N, Harmati G, Magyar J, Marangoni S, Zaza A, Varro A & Nanasi PP (2009). Reverse rate dependency is an intrinsic property of canine cardiac preparations. Cardiovasc Res 84, 237–244. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical