

Wnt and Neuregulin1/ErbB signalling extends 3D culture of hormone responsive mammary organoids
- PMID: 27782124
- PMCID: PMC5095178
- DOI: 10.1038/ncomms13207
Wnt and Neuregulin1/ErbB signalling extends 3D culture of hormone responsive mammary organoids
Abstract
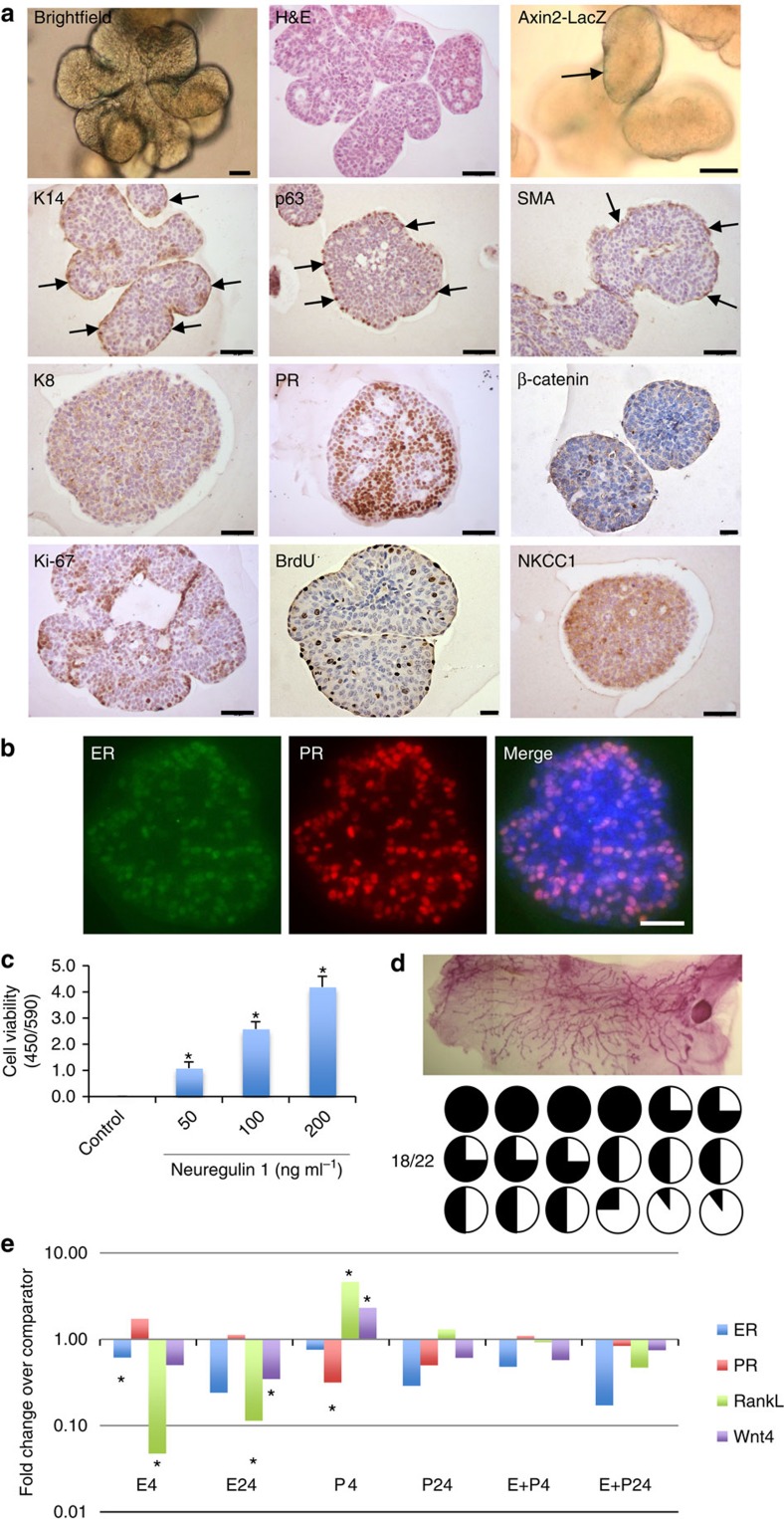
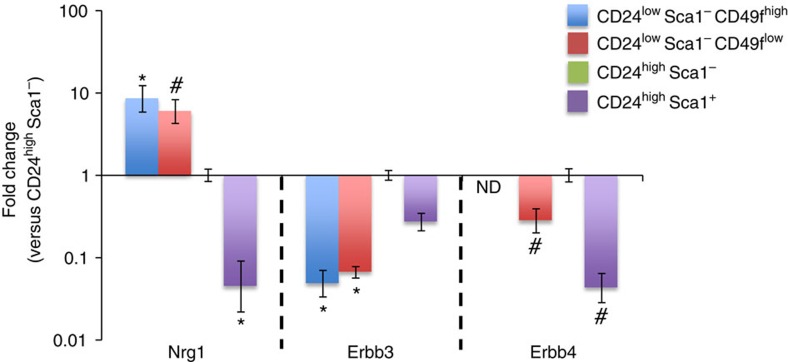
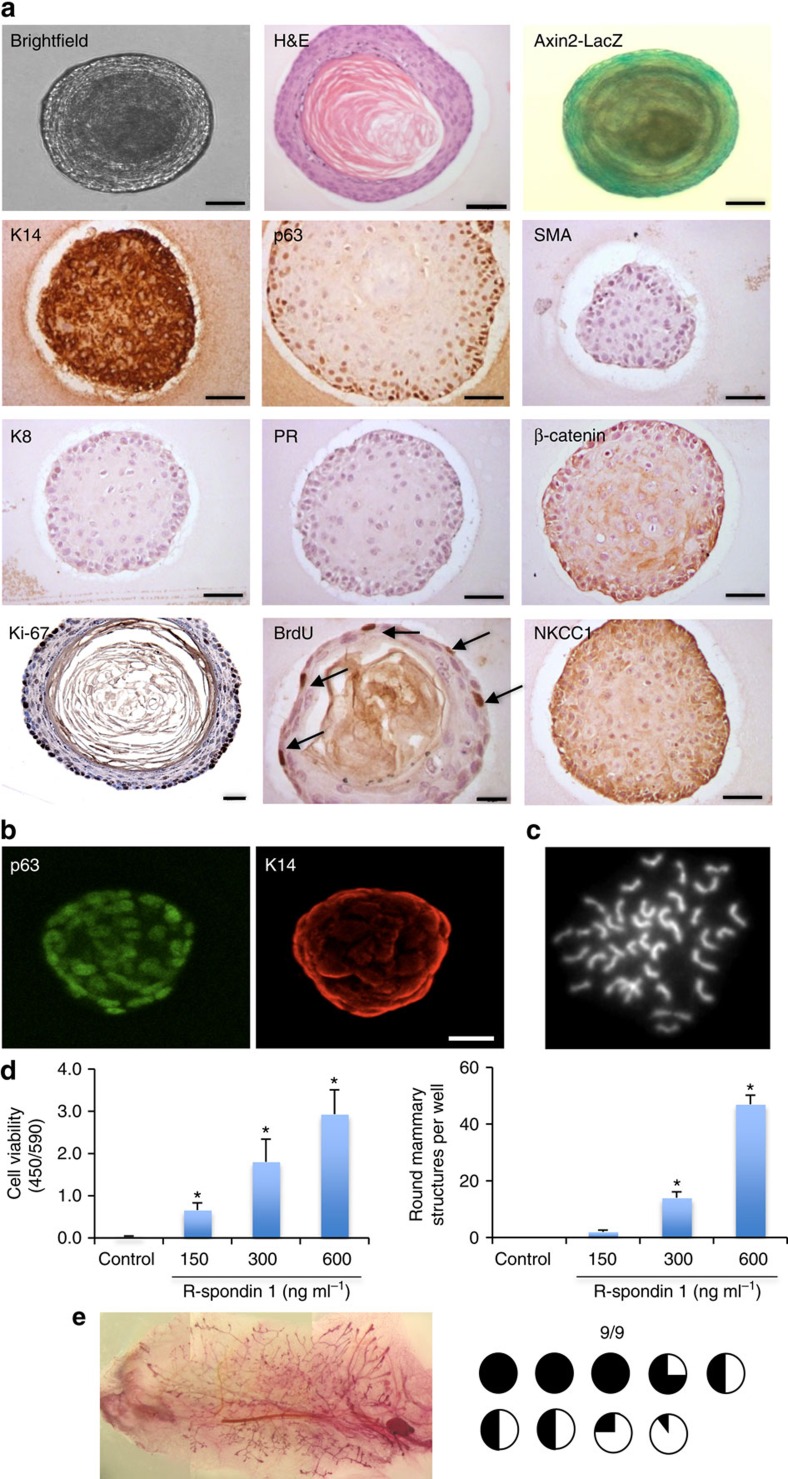
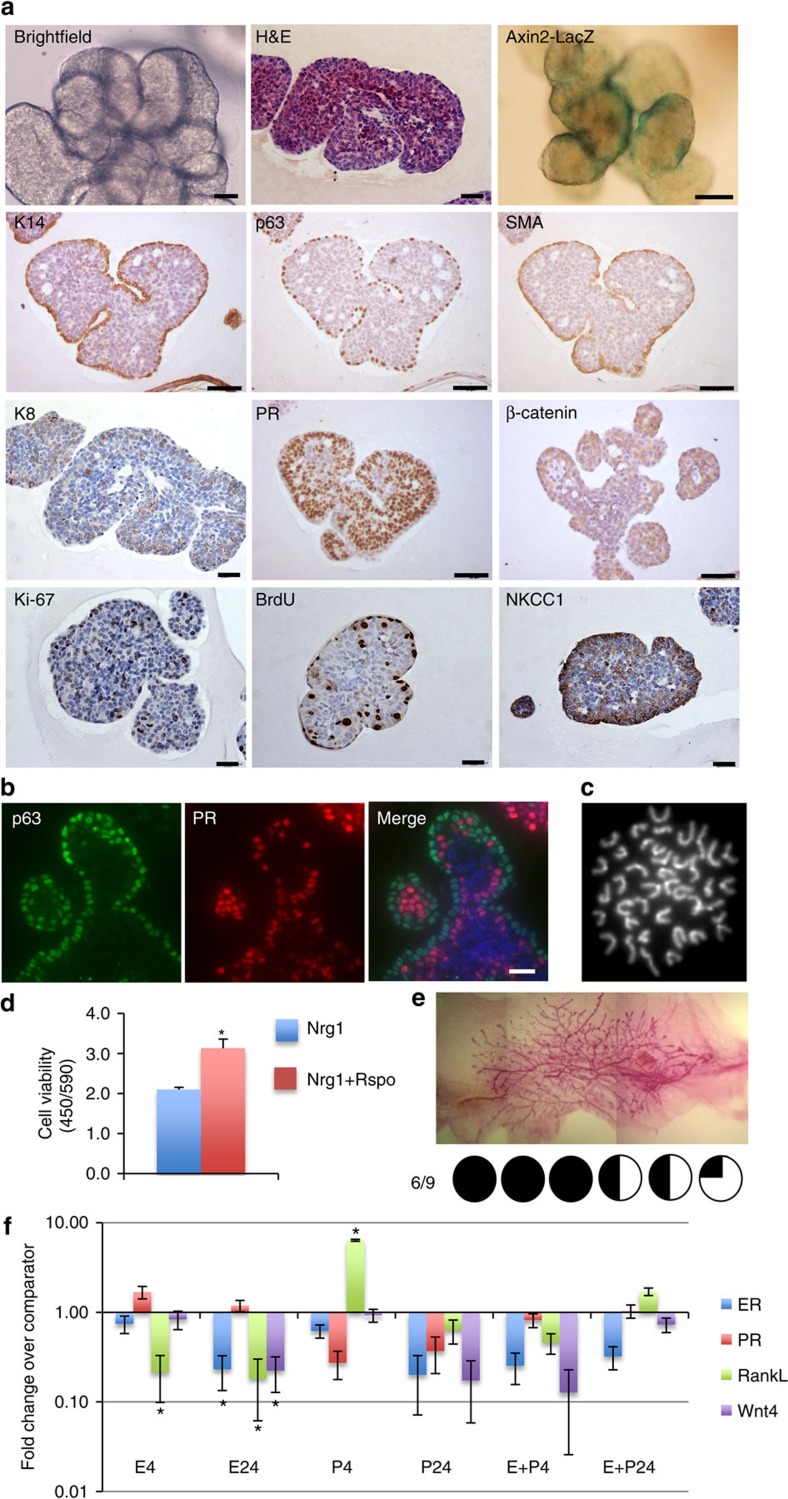
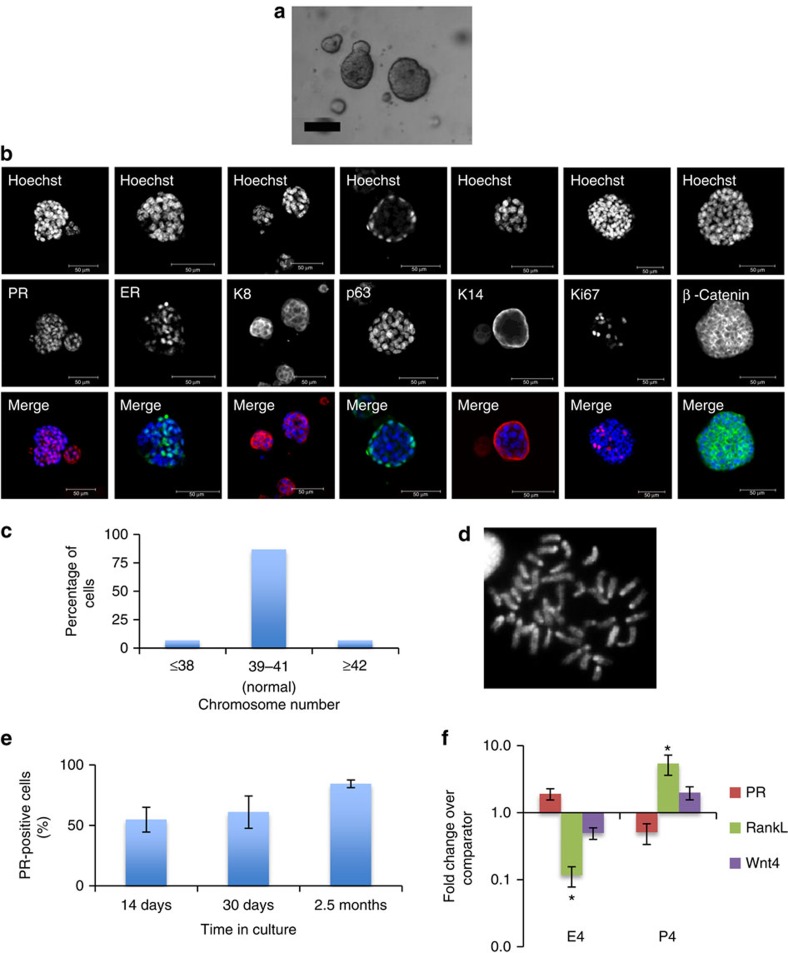
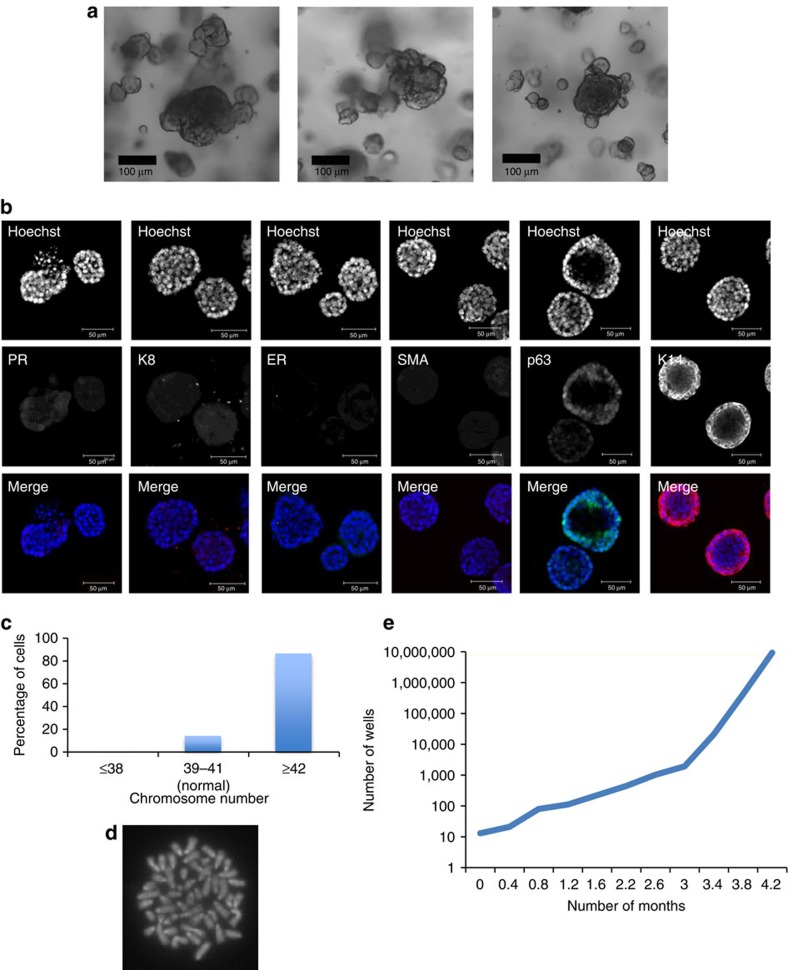
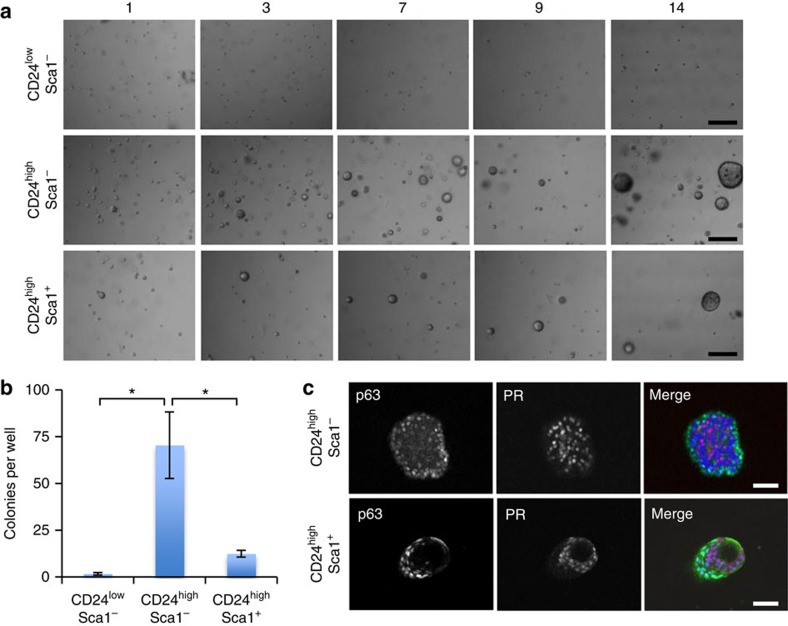
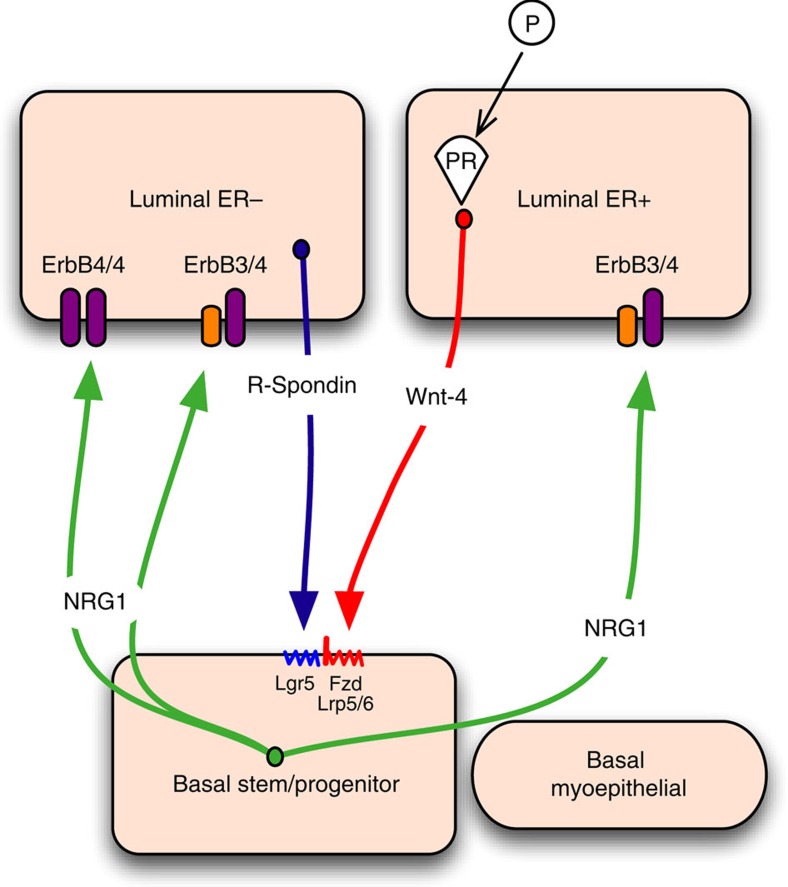
The development of in vitro culture systems quantitatively and qualitatively recapitulating normal breast biology is key to the understanding of mammary gland biology. Current three-dimensional mammary culture systems have not demonstrated concurrent proliferation and functional differentiation ex vivo in any system for longer than 2 weeks. Here, we identify conditions including Neuregulin1 and R-spondin 1, allowing maintenance and expansion of mammary organoids for 2.5 months in culture. The organoids comprise distinct basal and luminal compartments complete with functional steroid receptors and stem/progenitor cells able to reconstitute a complete mammary gland in vivo. Alternative conditions are also described that promote enrichment of basal cells organized into multiple layers surrounding a keratinous core, reminiscent of structures observed in MMTV-Wnt1 tumours. These conditions comprise a unique tool that should further understanding of normal mammary gland development, the molecular mechanism of hormone action and signalling events whose deregulation leads to breast tumourigenesis.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
