

Peroxisome proliferator activated receptor gamma 2 modulates late pregnancy homeostatic metabolic adaptations
- PMID: 27782293
- PMCID: PMC5135083
- DOI: 10.2119/molmed.2015.00262
Peroxisome proliferator activated receptor gamma 2 modulates late pregnancy homeostatic metabolic adaptations
Abstract
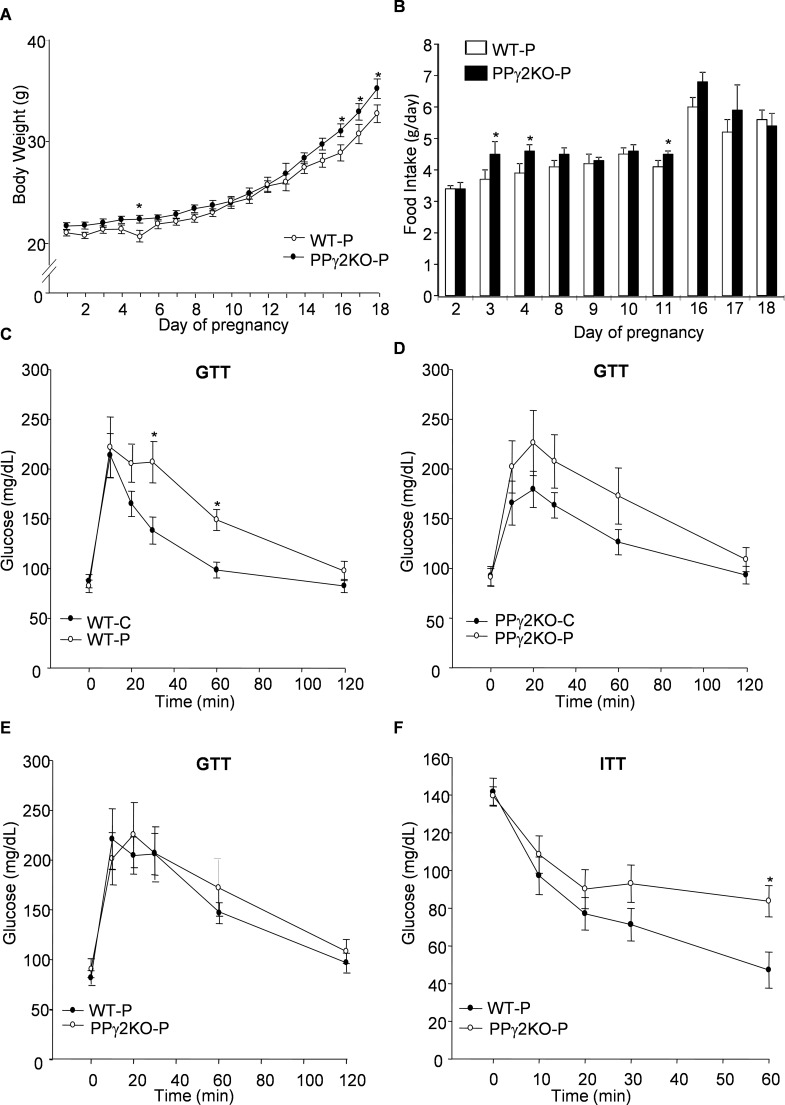
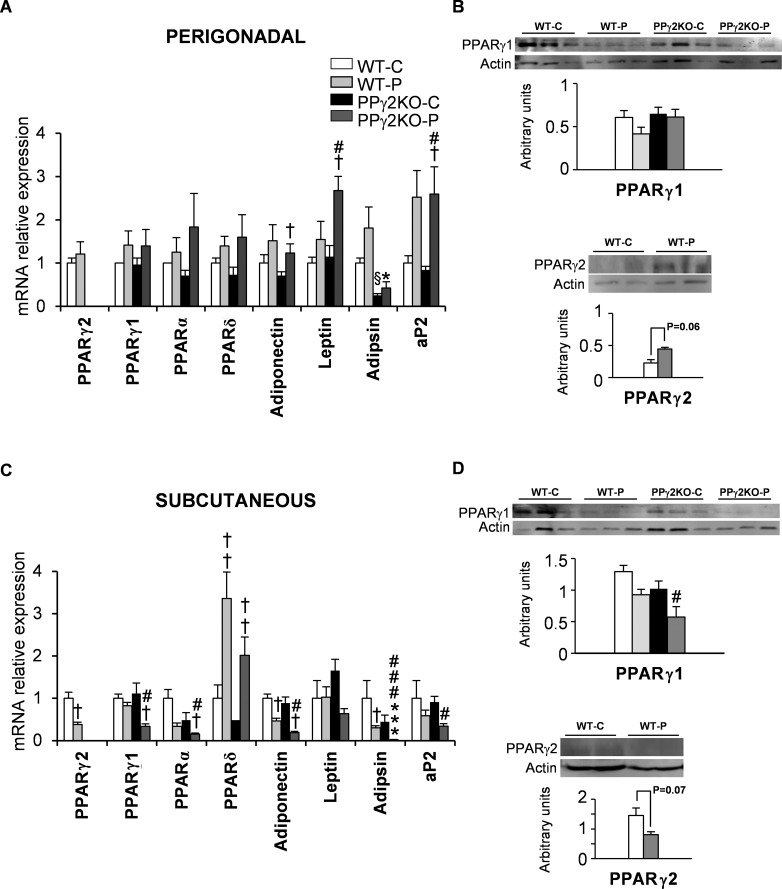
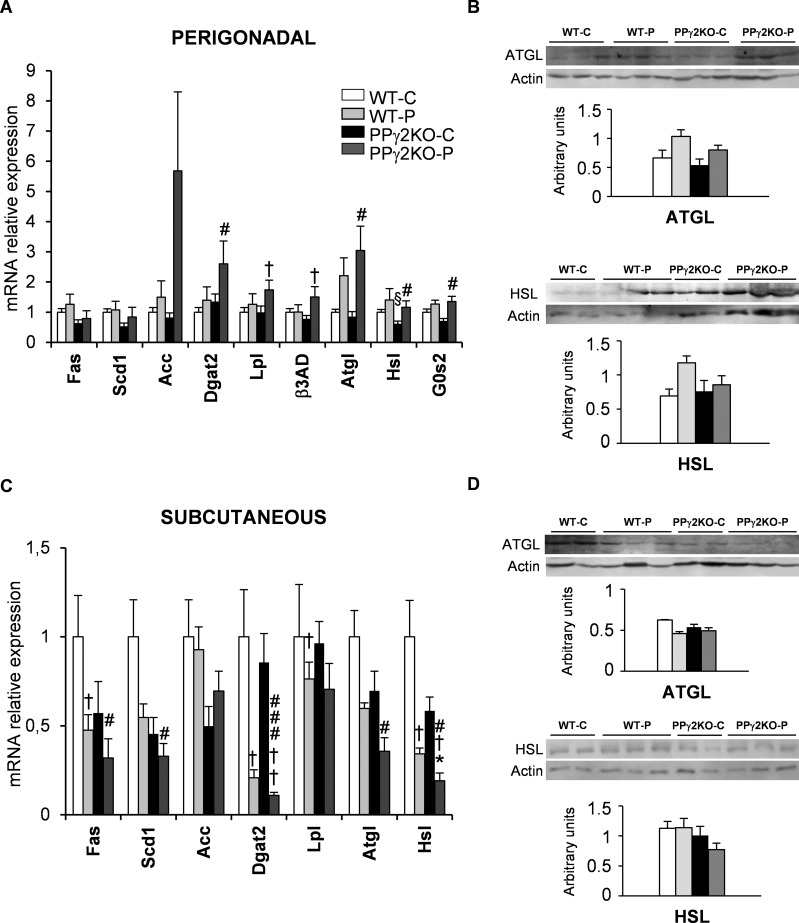
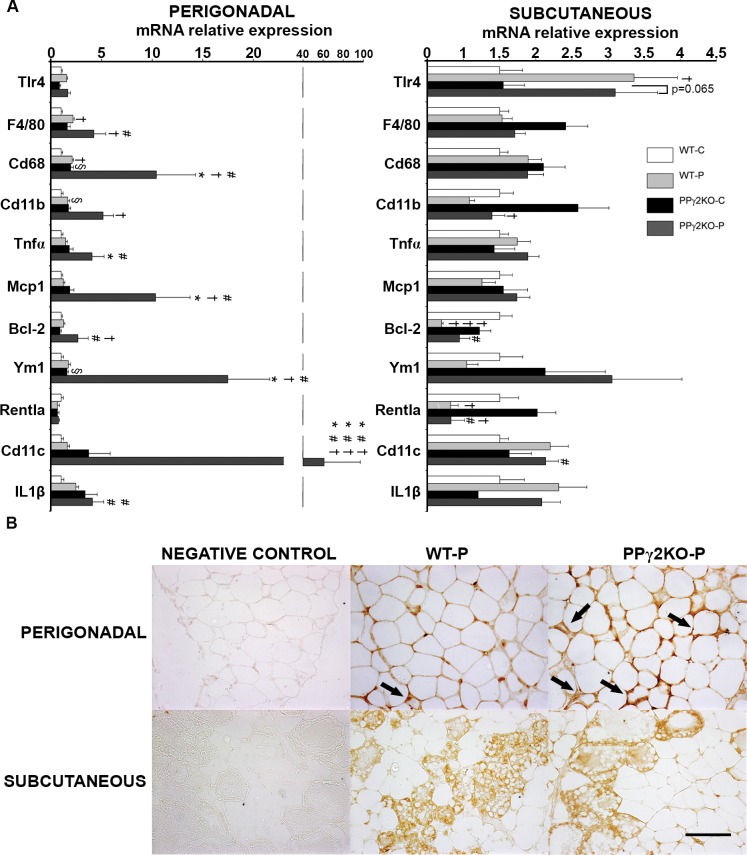
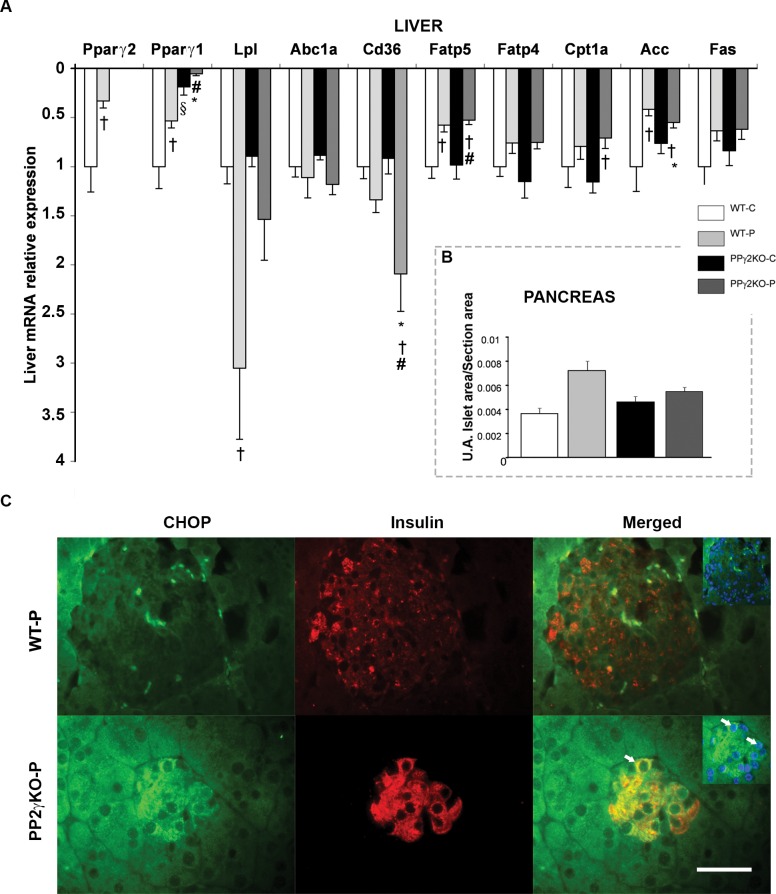
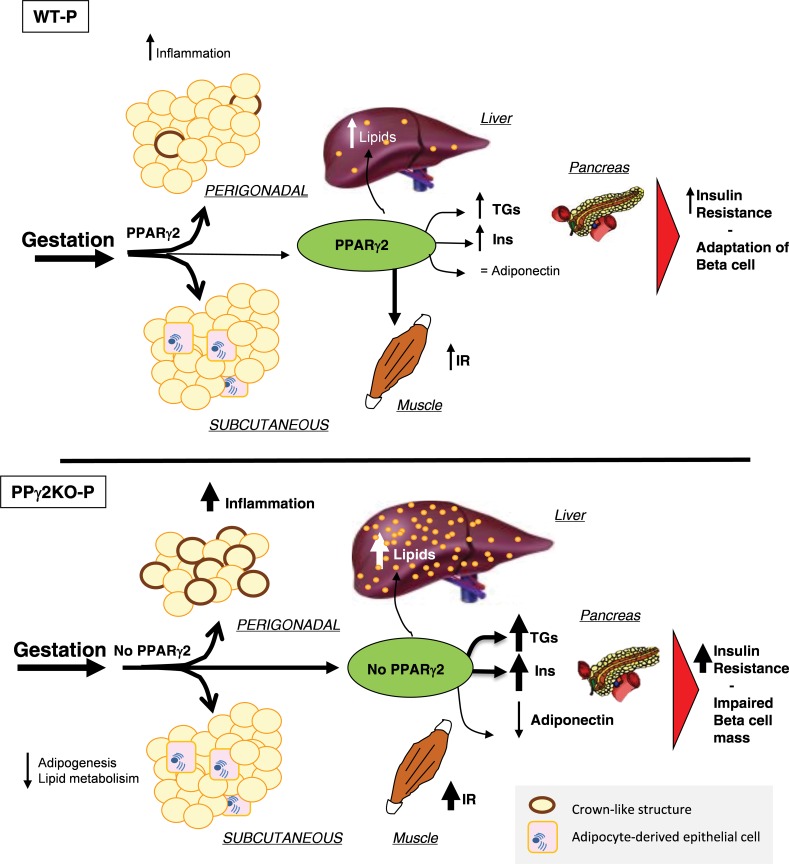
Pregnancy requires the adaptation of maternal energy metabolism including expansion and functional modifications of adipose tissue. Insulin resistance (IR), predominantly during late gestation, is a physiological metabolic adaptation that serves to support the metabolic demands of fetal growth. The molecular mechanisms underlying these adaptations are not fully understood and may contribute to gestational diabetes mellitus. Peroxisome proliferator-activated receptor gamma (PPARγ) controls adipogenesis, glucose and lipid metabolism and insulin sensitivity. The PPARγ2 isoform is mainly expressed in adipocytes and is thus likely to contribute to adipose tissue adaptation during late pregnancy. In the present study, we investigated the contribution of PPARγ2 to the metabolic adaptations occurring during the late phase of pregnancy in the context of IR. Using a model of late pregnancy in PPARγ2 knockout (KO) mice, we found that deletion of PPARγ2 exacerbated IR in association with lower serum adiponectin levels, increased body weight and enhanced lipid accumulation in liver. Lack of PPARγ2 provoked changes in the distribution of fat mass and preferentially prevented the expansion of the perigonadal depot while at the same time exacerbating inflammation. PPARγ2KO pregnant mice presented adipose tissue depot-dependent decreased expression of genes involved in lipid metabolism. Collectively, these data indicate that PPARγ2 is essential to promote healthy adipose tissue expansion and immune and metabolic functionality during pregnancy, contributing to the physiological adaptations that lead gestation to term.
Keywords: Insulin resistance; PPARγ; adiponectin; adipose tissue; diabetes; gestation; inflammation.
Conflict of interest statement
The authors declare they have no competing interests as defined by
Figures
References
-
- Ramos MP, Crespo-Solans MD, del Campo S, Cacho J, Herrera E. Fat accumulation in the rat during early pregnancy is modulated by enhanced insulin responsiveness. Am J Physiol Endocrinol Metab. 2003;285(2):E318–28. - PubMed
-
- Colomiere M, Permezel M, Lappas M. Diabetes and obesity during pregnancy alter insulin signalling and glucose transporter expression in maternal skeletal muscle and subcutaneous adipose tissue. J Mol Endocrinol. 2010;44(4):213–23. - PubMed
-
- Sevillano J, de Castro J, Bocos C, Herrera E, Ramos MP. Role of insulin receptor substrate-1 serine 307 phosphorylation and adiponectin in adipose tissue insulin resistance in late pregnancy. Endocrinology. 2007;148(12):5933–42. - PubMed
-
- Barbarroja N, et al. The obese healthy paradox: is inflammation the answer? Biochem J. 2010;430(1):141–49. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
