

Roles of H3K27me2 and H3K27me3 Examined during Fate Specification of Embryonic Stem Cells
- PMID: 27783950
- PMCID: PMC5123747
- DOI: 10.1016/j.celrep.2016.09.087
Roles of H3K27me2 and H3K27me3 Examined during Fate Specification of Embryonic Stem Cells
Erratum in
-
Roles of H3K27me2 and H3K27me3 Examined during Fate Specification of Embryonic Stem Cells.Cell Rep. 2017 Jan 3;18(1):297. doi: 10.1016/j.celrep.2016.12.036. Cell Rep. 2017. PMID: 28052258 No abstract available.
Abstract
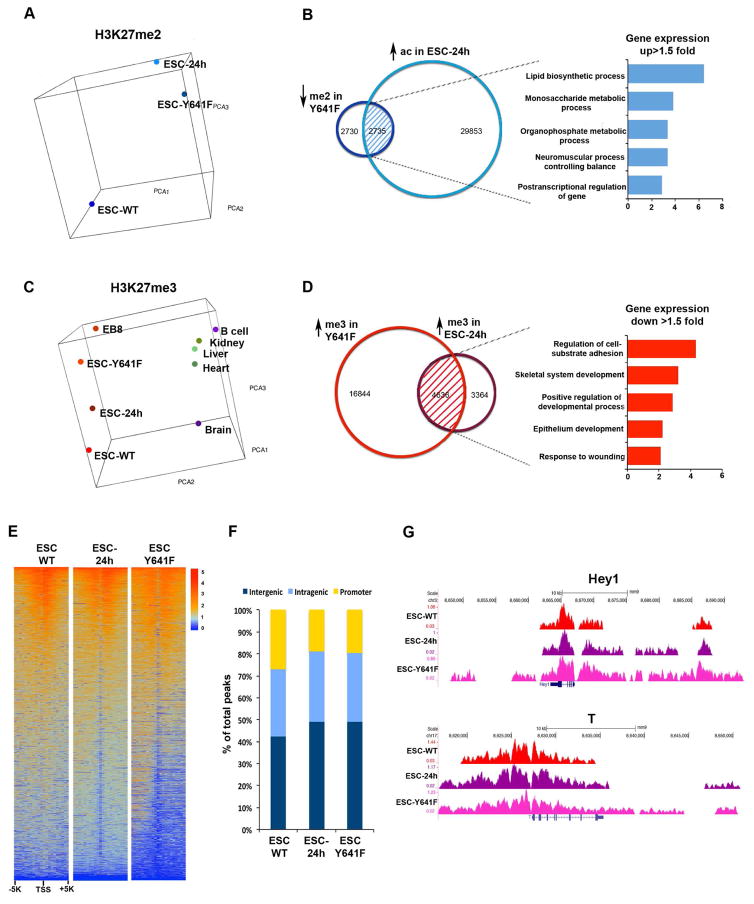
The polycomb repressive complex 2 (PRC2) methylates lysine 27 of histone H3 (H3K27) through its catalytic subunit Ezh2. PRC2-mediated di- and tri-methylation (H3K27me2/H3K27me3) have been interchangeably associated with gene repression. However, it remains unclear whether these two degrees of H3K27 methylation have different functions. In this study, we have generated isogenic mouse embryonic stem cells (ESCs) with a modified H3K27me2/H3K27me3 ratio. Our findings document dynamic developmental control in the genomic distribution of H3K27me2 and H3K27me3 at regulatory regions in ESCs. They also reveal that modifying the ratio of H3K27me2 and H3K27me3 is sufficient for the acquisition and repression of defined cell lineage transcriptional programs and phenotypes and influences induction of the ESC ground state.
Keywords: H3K27 methylation; embryonic stem cells; polycomb proteins.
Published by Elsevier Inc.
Figures





References
-
- Azuara V, Perry P, Sauer S, Spivakov M, Jorgensen HF, John RM, Gouti M, Casanova M, Warnes G, Merkenschlager M, et al. Chromatin signatures of pluripotent cell lines. Nature cell biology. 2006;8:532–538. - PubMed
-
- Barsotti AM, Ryskin M, Zhong W, Zhang WG, Giannakou A, Loreth C, Diesl V, Follettie M, Golas J, Lee M, et al. Epigenetic reprogramming by tumor-derived EZH2 gain-of-function mutations promotes aggressive 3D cell morphologies and enhances melanoma tumor growth. Oncotarget. 2015;6:2928–2938. - PMC - PubMed
-
- Ben-Zvi D, Melton DA. Modeling human nutrition using human embryonic stem cells. Cell. 2015;161:12–17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
