

The fungal-specific β-glucan-binding lectin FGB1 alters cell-wall composition and suppresses glucan-triggered immunity in plants
- PMID: 27786272
- PMCID: PMC5095285
- DOI: 10.1038/ncomms13188
The fungal-specific β-glucan-binding lectin FGB1 alters cell-wall composition and suppresses glucan-triggered immunity in plants
Abstract
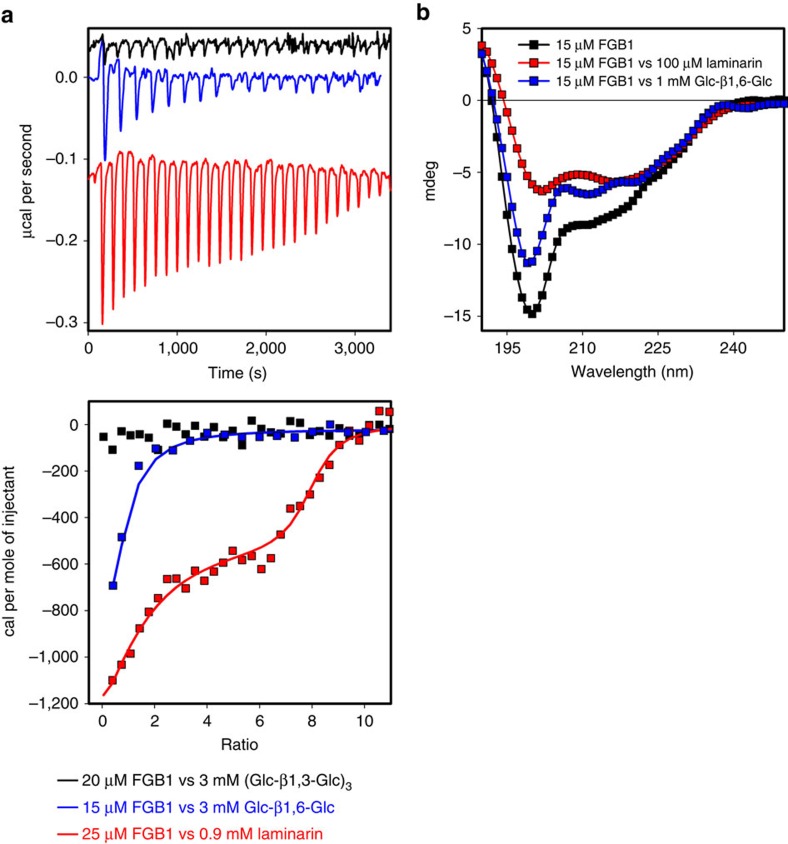
β-glucans are well-known modulators of the immune system in mammals but little is known about β-glucan triggered immunity in planta. Here we show by isothermal titration calorimetry, circular dichroism spectroscopy and nuclear magnetic resonance spectroscopy that the FGB1 gene from the root endophyte Piriformospora indica encodes for a secreted fungal-specific β-glucan-binding lectin with dual function. This lectin has the potential to both alter fungal cell wall composition and properties, and to efficiently suppress β-glucan-triggered immunity in different plant hosts, such as Arabidopsis, barley and Nicotiana benthamiana. Our results hint at the existence of fungal effectors that deregulate innate sensing of β-glucan in plants.
Figures




References
-
- Spoel S. H. & Dong X. N. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 12, 89–100 (2012). - PubMed
-
- Jones J. D. & Dangl J. L. The plant immune system. Nature 444, 323–329 (2006). - PubMed
-
- Limpens E., van Zeijl A. & Geurts R. Lipochitooligosaccharides modulate plant host immunity to enable endosymbioses. Annu. Rev. Phytopathol. 53, 311–334 (2015). - PubMed
-
- Nurnberger T., Brunner F., Kemmerling B. & Piater L. Innate immunity in plants and animals: striking similarities and obvious differences. Immunol. Rev. 198, 249–266 (2004). - PubMed
-
- Sanchez-Vallet A., Mesters J. R. & Thomma B. P. The battle for chitin recognition in plant-microbe interactions. FEMS Microbiol. Rev. 39, 171–183 (2015). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
