

Canonical Wnt signalling regulates nuclear export of Setdb1 during skeletal muscle terminal differentiation
- PMID: 27790377
- PMCID: PMC5067623
- DOI: 10.1038/celldisc.2016.37
Canonical Wnt signalling regulates nuclear export of Setdb1 during skeletal muscle terminal differentiation
Erratum in
-
Erratum: Canonical Wnt signalling regulates nuclear export of Setdb1 during skeletal muscle terminal differentiation.Cell Discov. 2016 Nov 15;2:16043. doi: 10.1038/celldisc.2016.43. eCollection 2016. Cell Discov. 2016. PMID: 27867535 Free PMC article.
Abstract
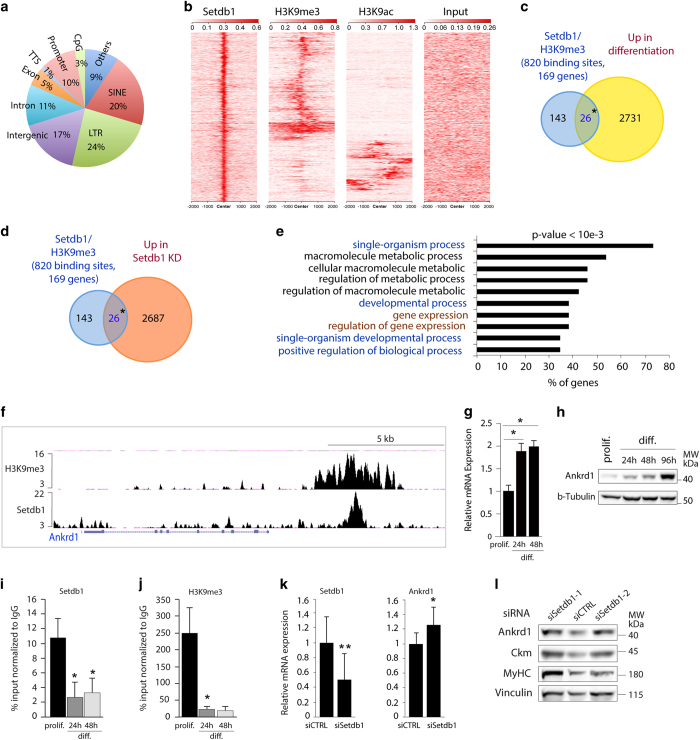
The histone 3 lysine 9 methyltransferase Setdb1 is essential for both stem cell pluripotency and terminal differentiation of different cell types. To shed light on the roles of Setdb1 in these mutually exclusive processes, we used mouse skeletal myoblasts as a model of terminal differentiation. Ex vivo studies on isolated single myofibres showed that Setdb1 is required for adult muscle stem cells expansion following activation. In vitro studies in skeletal myoblasts confirmed that Setdb1 suppresses terminal differentiation. Genomic binding analyses showed a release of Setdb1 from selected target genes upon myoblast terminal differentiation, concomitant to a nuclear export of Setdb1 to the cytoplasm. Both genomic release and cytoplasmic Setdb1 relocalisation during differentiation were dependent on canonical Wnt signalling. Transcriptomic assays in myoblasts unravelled a significant overlap between Setdb1 and Wnt3a regulated genetic programmes. Together, our findings revealed Wnt-dependent subcellular relocalisation of Setdb1 as a novel mechanism regulating Setdb1 functions and myogenesis.
Keywords: SETDB1/ESET/KMT1E; Wnt signalling; differentiation; lysine; methyltransferase; muscle.
Figures





References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
