

Context-specific inhibition of translation by ribosomal antibiotics targeting the peptidyl transferase center
- PMID: 27791002
- PMCID: PMC5086994
- DOI: 10.1073/pnas.1613055113
Context-specific inhibition of translation by ribosomal antibiotics targeting the peptidyl transferase center
Abstract
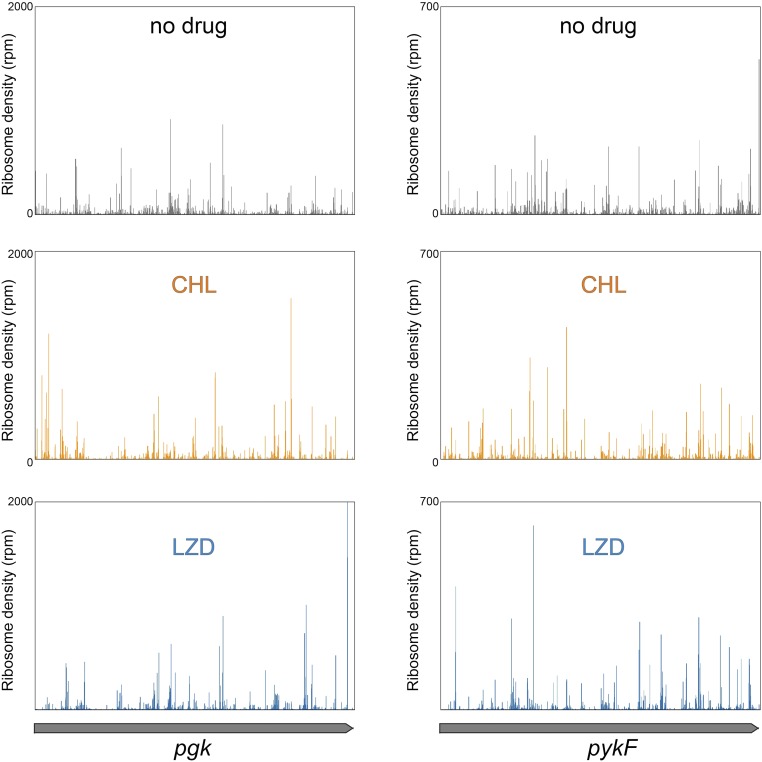
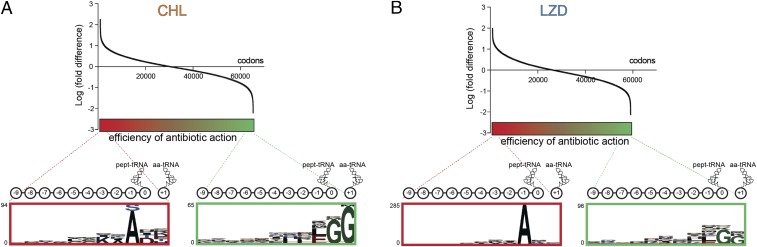
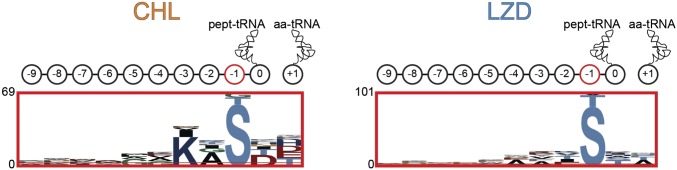
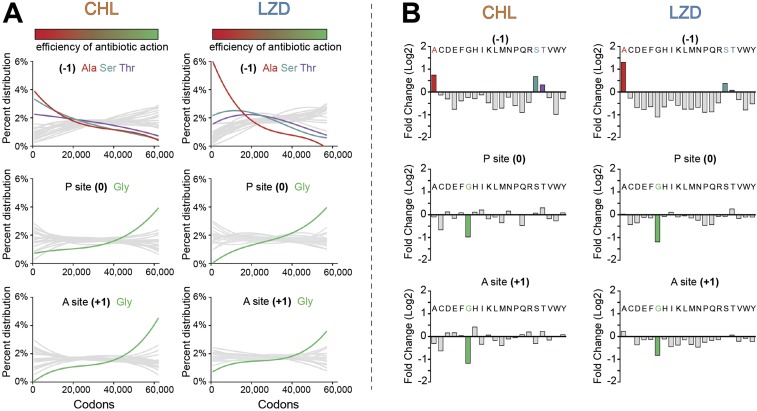
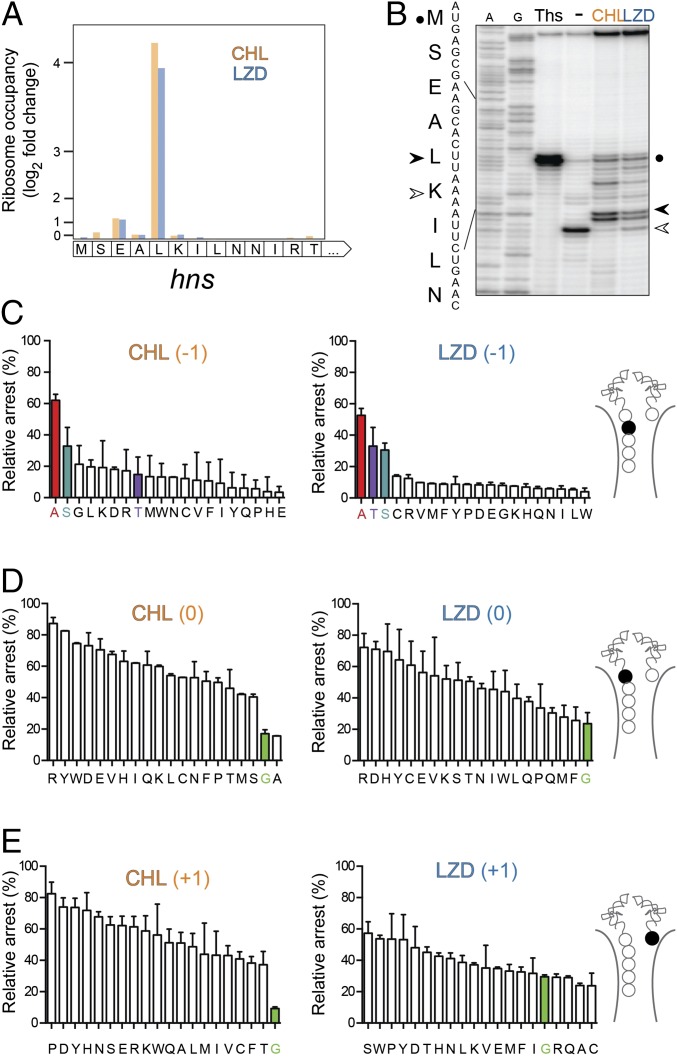
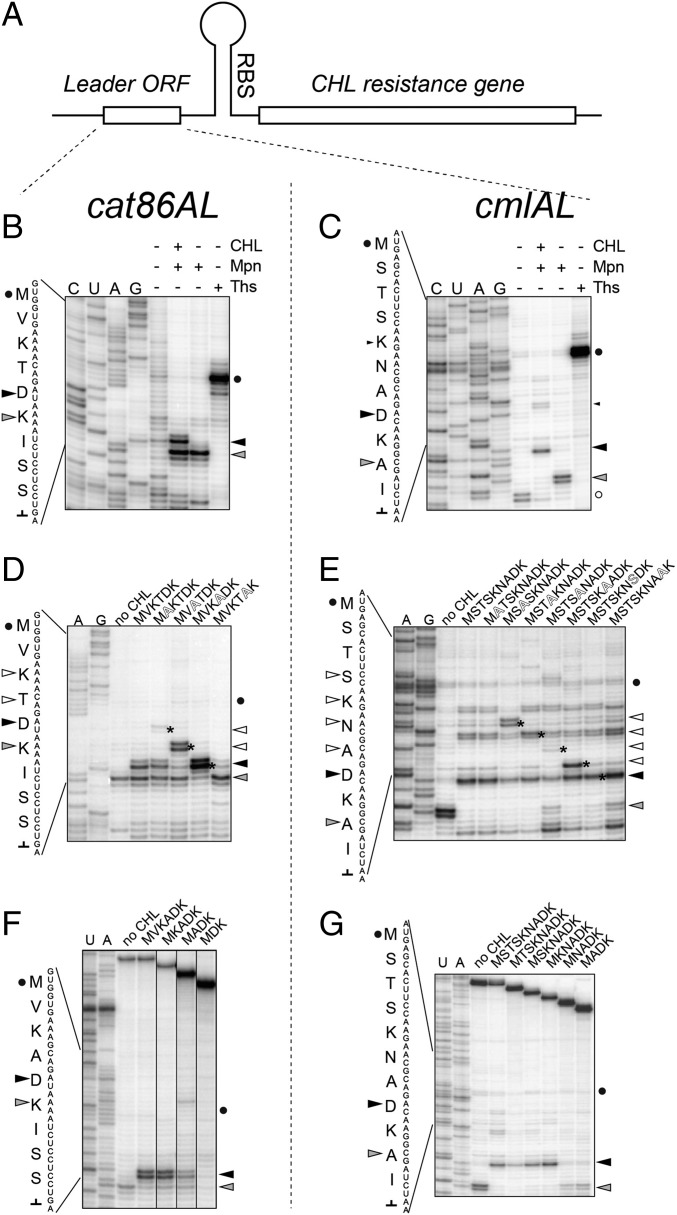
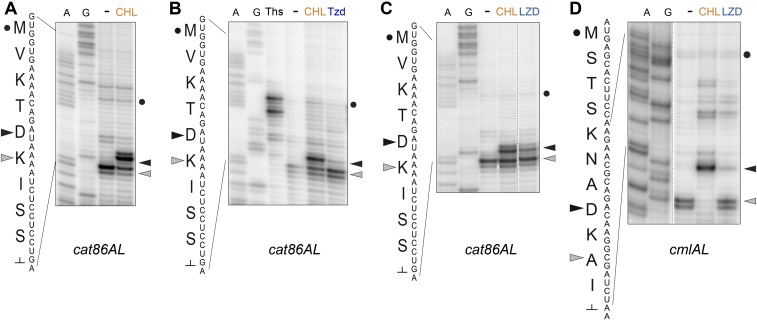
The first broad-spectrum antibiotic chloramphenicol and one of the newest clinically important antibacterials, linezolid, inhibit protein synthesis by targeting the peptidyl transferase center of the bacterial ribosome. Because antibiotic binding should prevent the placement of aminoacyl-tRNA in the catalytic site, it is commonly assumed that these drugs are universal inhibitors of peptidyl transfer and should readily block the formation of every peptide bond. However, our in vitro experiments showed that chloramphenicol and linezolid stall ribosomes at specific mRNA locations. Treatment of bacterial cells with high concentrations of these antibiotics leads to preferential arrest of translation at defined sites, resulting in redistribution of the ribosomes on mRNA. Antibiotic-mediated inhibition of protein synthesis is most efficient when the nascent peptide in the ribosome carries an alanine residue and, to a lesser extent, serine or threonine in its penultimate position. In contrast, the inhibitory action of the drugs is counteracted by glycine when it is either at the nascent-chain C terminus or at the incoming aminoacyl-tRNA. The context-specific action of chloramphenicol illuminates the operation of the mechanism of inducible resistance that relies on programmed drug-induced translation arrest. In addition, our findings expose the functional interplay between the nascent chain and the peptidyl transferase center.
Keywords: antibiotics; nascent peptide; oxazolidinones; protein synthesis; ribosome.
Conflict of interest statement
The research in the laboratory was supported by, among other sources, grants from the pharmaceutical companies Melinta Therapeutics and Cempra Pharmaceuticals.
Figures





Comment in
-
Antimicrobials: Putting antibiotic action into context.Nat Rev Microbiol. 2016 Dec;14(12):725. doi: 10.1038/nrmicro.2016.165. Epub 2016 Oct 31. Nat Rev Microbiol. 2016. PMID: 27795566 No abstract available.
References
-
- Beringer M, Rodnina MV. The ribosomal peptidyl transferase. Mol Cell. 2007;26(3):311–321. - PubMed
-
- Weinger JS, Parnell KM, Dorner S, Green R, Strobel SA. Substrate-assisted catalysis of peptide bond formation by the ribosome. Nat Struct Mol Biol. 2004;11(11):1101–1106. - PubMed
-
- Polacek N, Gaynor M, Yassin A, Mankin AS. Ribosomal peptidyl transferase can withstand mutations at the putative catalytic nucleotide. Nature. 2001;411(6836):498–501. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
