

Notch3 drives development and progression of cholangiocarcinoma
- PMID: 27791012
- PMCID: PMC5086988
- DOI: 10.1073/pnas.1600067113
Notch3 drives development and progression of cholangiocarcinoma
Abstract
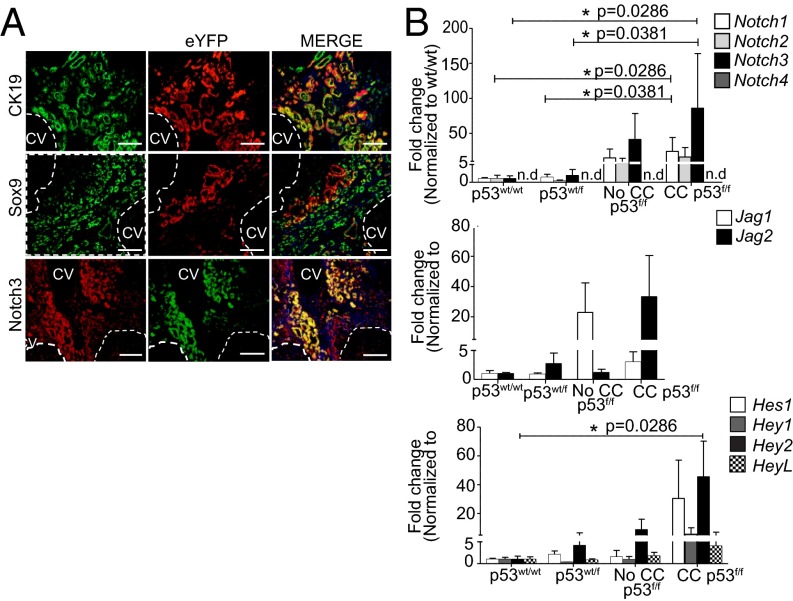
The prognosis of cholangiocarcinoma (CC) is dismal. Notch has been identified as a potential driver; forced exogenous overexpression of Notch1 in hepatocytes results in the formation of biliary tumors. In human disease, however, it is unknown which components of the endogenously signaling pathway are required for tumorigenesis, how these orchestrate cancer, and how they can be targeted for therapy. Here we characterize Notch in human-resected CC, a toxin-driven model in rats, and a transgenic mouse model in which p53 deletion is targeted to biliary epithelia and CC induced using the hepatocarcinogen thioacetamide. We find that across species, the atypical receptor NOTCH3 is differentially overexpressed; it is progressively up-regulated with disease development and promotes tumor cell survival via activation of PI3k-Akt. We use genetic KO studies to show that tumor growth significantly attenuates after Notch3 deletion and demonstrate signaling occurs via a noncanonical pathway independent of the mediator of classical Notch, Recombinant Signal Binding Protein for Immunoglobulin Kappa J Region (RBPJ). These data present an opportunity in this aggressive cancer to selectively target Notch, bypassing toxicities known to be RBPJ dependent.
Keywords: Notch; bile duct; cancer; cholangiocarcinoma; noncanonical.
Conflict of interest statement
The authors declare no conflict of interest.
Figures













References
-
- Ortica S, Tarantino N, Aulner N, Israel A, Gupta-Rossi N. The 4 Notch receptors play distinct and antagonistic roles in the proliferation and hepatocytic differentiation of liver progenitors. FASEB J. 2014;28(2):603–614. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
