

Increasing seed size and quality by manipulating BIG SEEDS1 in legume species
- PMID: 27791139
- PMCID: PMC5098654
- DOI: 10.1073/pnas.1611763113
Increasing seed size and quality by manipulating BIG SEEDS1 in legume species
Abstract
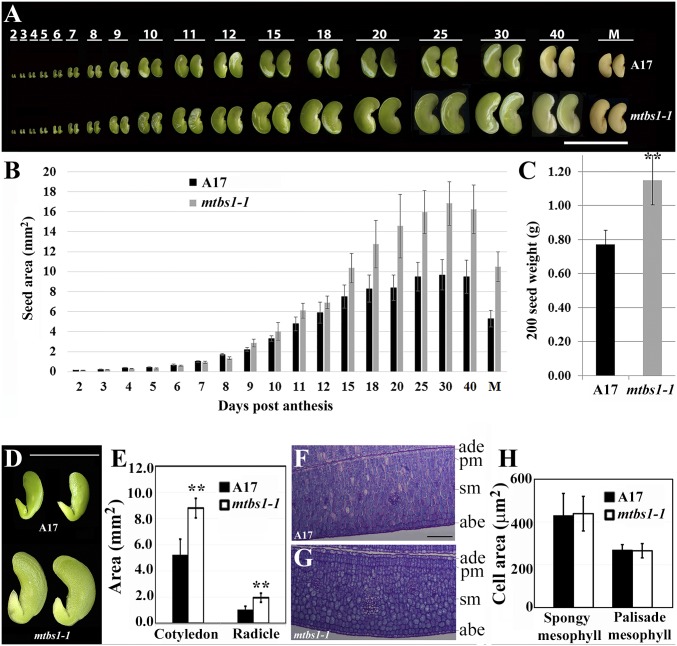
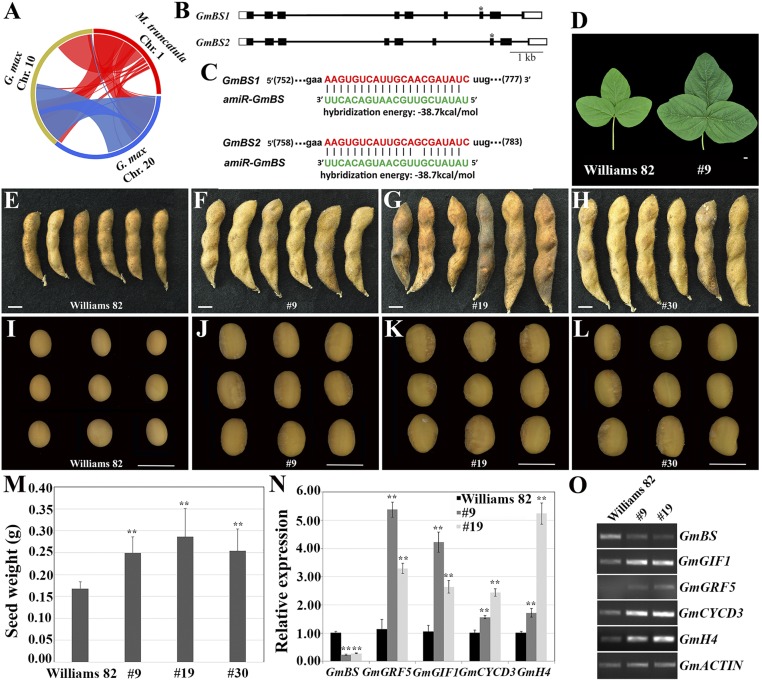
Plant organs, such as seeds, are primary sources of food for both humans and animals. Seed size is one of the major agronomic traits that have been selected in crop plants during their domestication. Legume seeds are a major source of dietary proteins and oils. Here, we report a conserved role for the BIG SEEDS1 (BS1) gene in the control of seed size and weight in the model legume Medicago truncatula and the grain legume soybean (Glycine max). BS1 encodes a plant-specific transcription regulator and plays a key role in the control of the size of plant organs, including seeds, seed pods, and leaves, through a regulatory module that targets primary cell proliferation. Importantly, down-regulation of BS1 orthologs in soybean by an artificial microRNA significantly increased soybean seed size, weight, and amino acid content. Our results provide a strategy for the increase in yield and seed quality in legumes.
Keywords: Medicago; forage quality; plant organ size; seed size; soybean.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Lam HM, et al. Resequencing of 31 wild and cultivated soybean genomes identifies patterns of genetic diversity and selection. Nat Genet. 2010;42(12):1053–1059. - PubMed
-
- Schmutz J, et al. Genome sequence of the palaeopolyploid soybean. Nature. 2010;463(7278):178–183. - PubMed
-
- Masuda T, Goldsmith PD. World soybean production: Area harvested, yield, and long-term projections. Int Food Agribusiness Manage Rev. 2009;12(4):143–162.
-
- Linkies A, Graeber K, Knight C, Leubner-Metzger G. The evolution of seeds. New Phytol. 2010;186(4):817–831. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
