

p62- and ubiquitin-dependent stress-induced autophagy of the mammalian 26S proteasome
- PMID: 27791183
- PMCID: PMC5127335
- DOI: 10.1073/pnas.1615455113
p62- and ubiquitin-dependent stress-induced autophagy of the mammalian 26S proteasome
Abstract
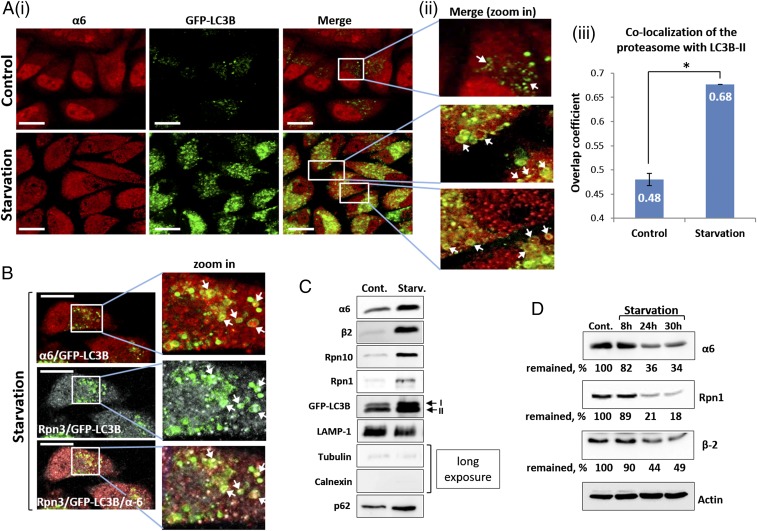
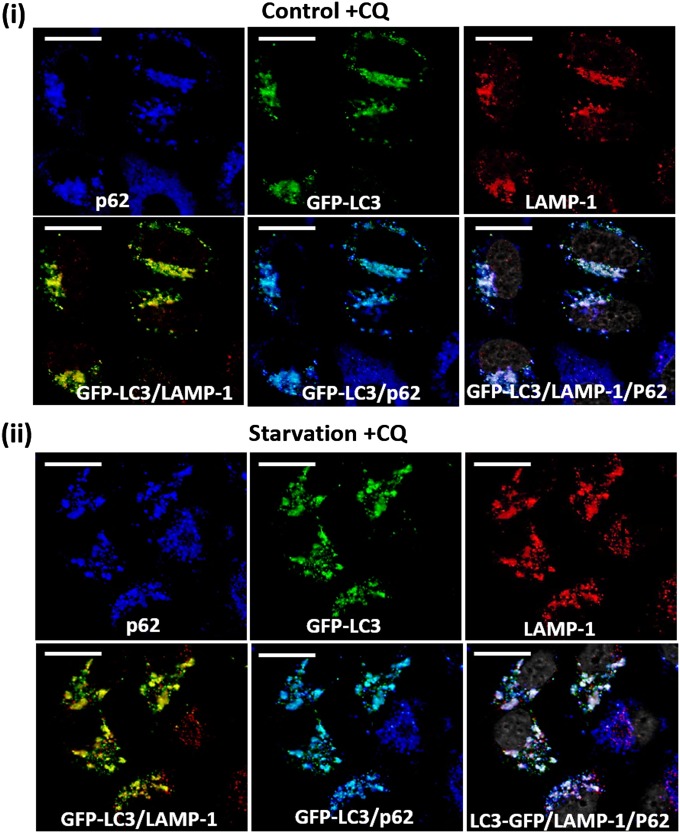
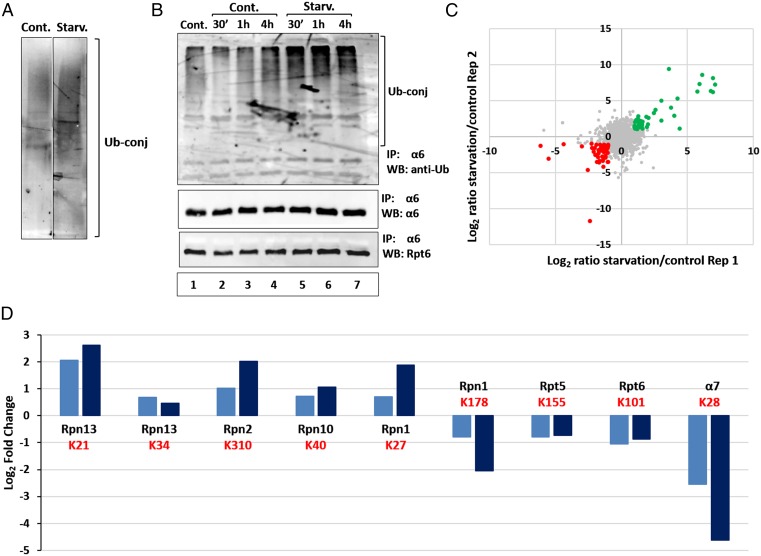
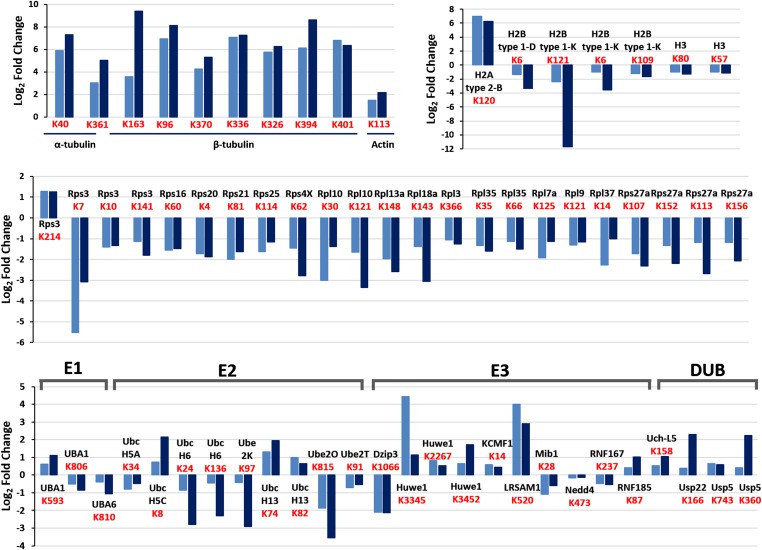
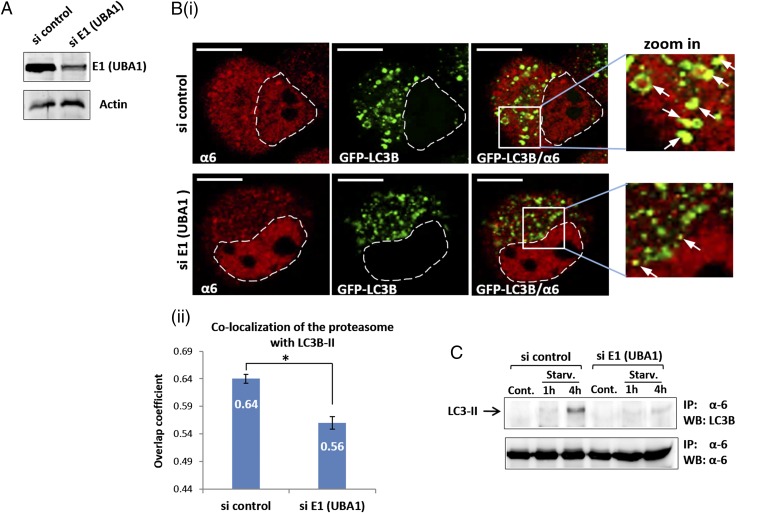
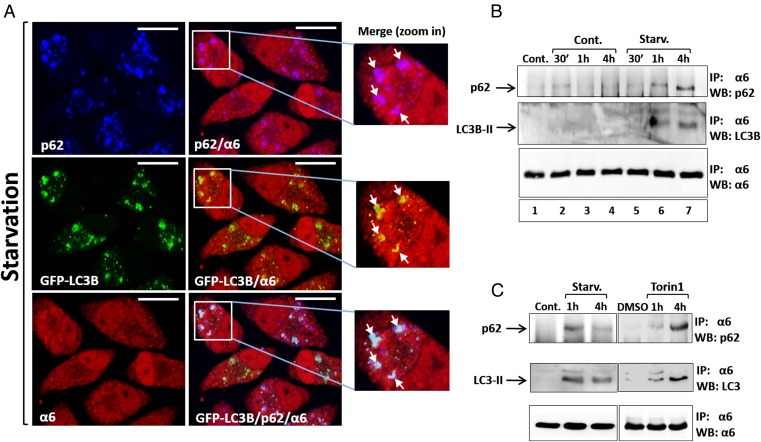
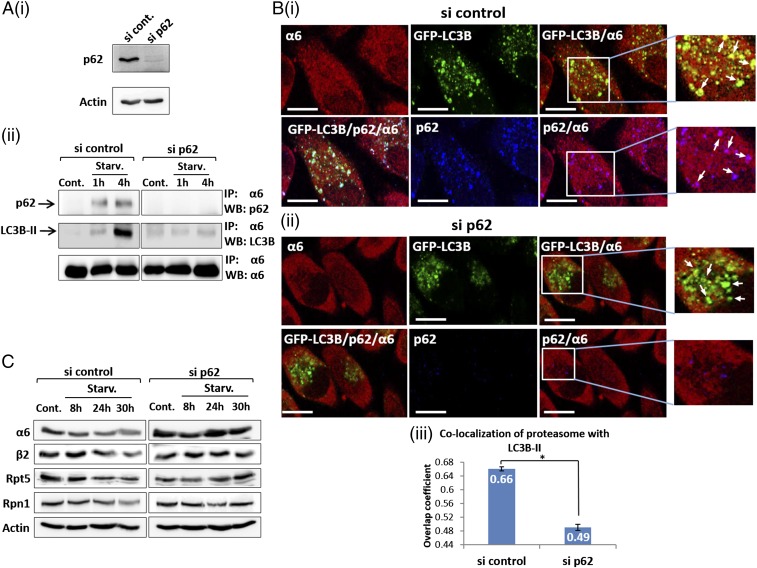
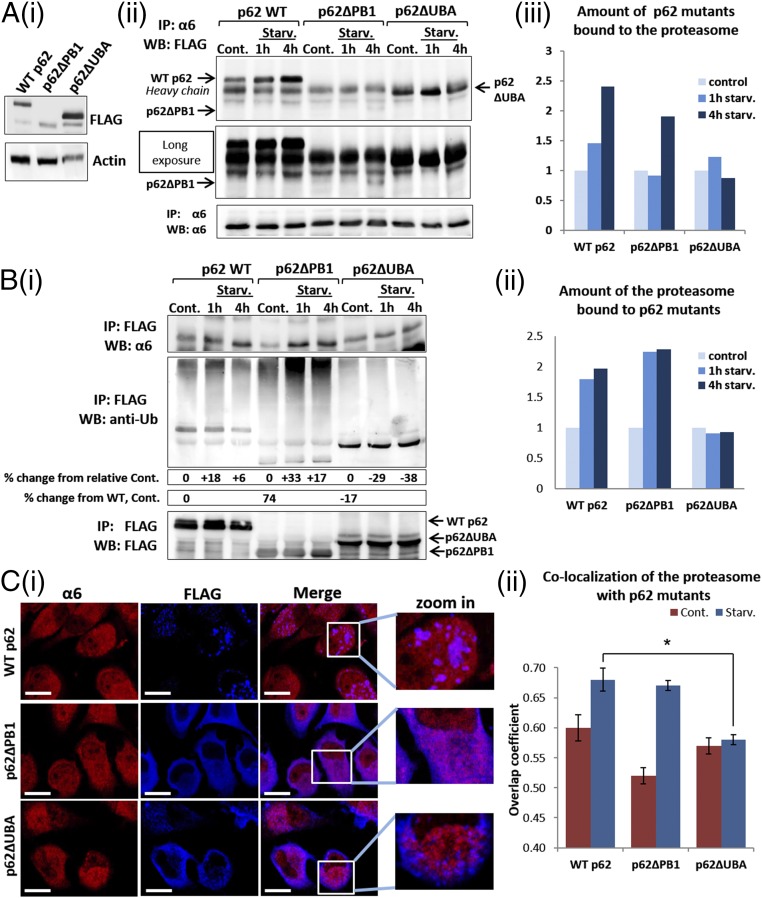
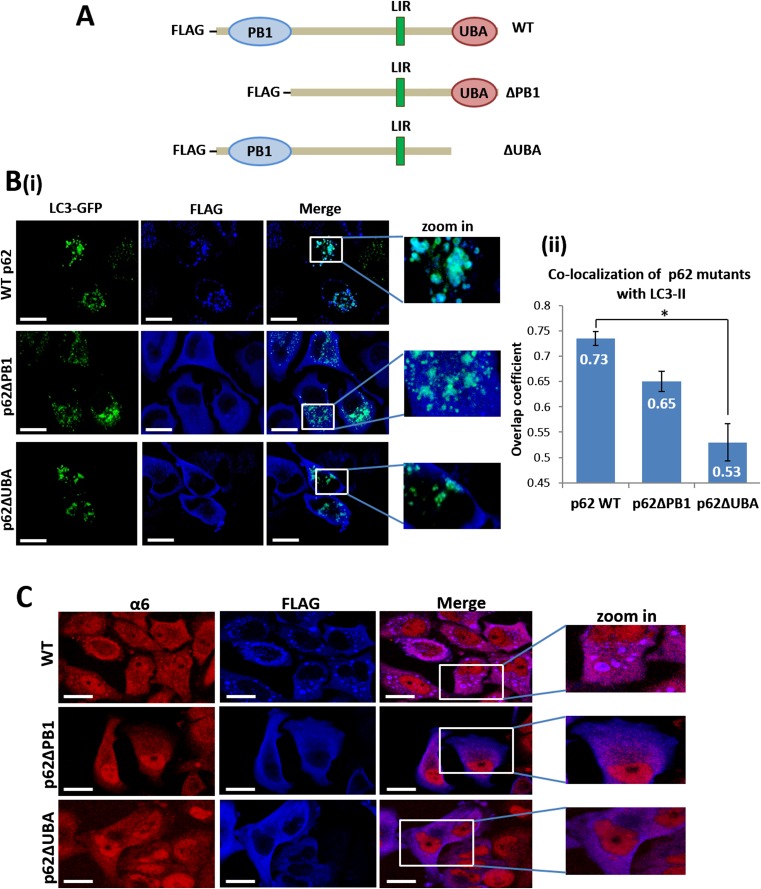
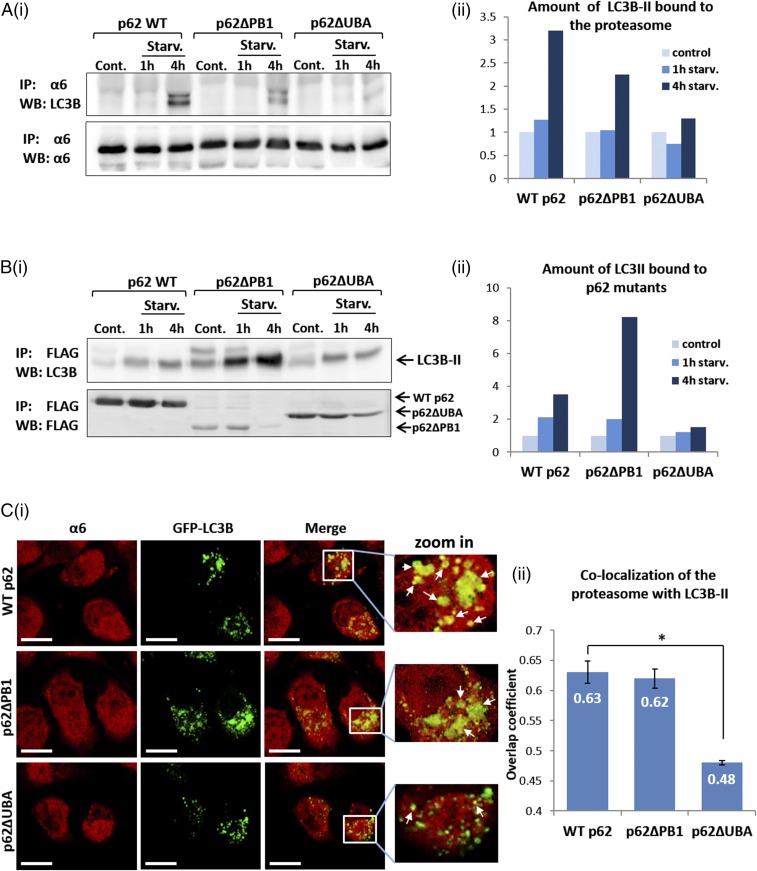
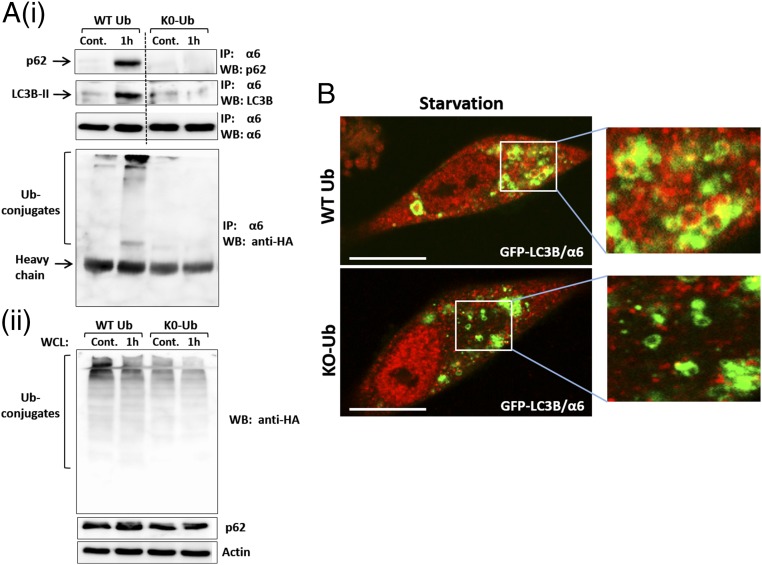
The ubiquitin-proteasome system and autophagy are the two main proteolytic systems involved in, among other functions, the maintenance of cell integrity by eliminating misfolded and damaged proteins and organelles. Both systems remove their targets after their conjugation with ubiquitin. An interesting, yet incompletely understood problem relates to the fate of the components of the two systems. Here we provide evidence that amino acid starvation enhances polyubiquitination on specific sites of the proteasome, a modification essential for its targeting to the autophagic machinery. The uptake of the ubiquitinated proteasome is mediated by its interaction with the ubiquitin-associated domain of p62/SQSTM1, a process that also requires interaction with LC3. Importantly, deletion of the PB1 domain of p62, which is important for the targeting of ubiquitinated substrates to the proteasome, has no effect on stress-induced autophagy of this proteolytic machinery, suggesting that the domain of p62 that binds to the proteasome determines the function of p62 in either targeting substrates to the proteasome or targeting the proteasome to autophagy.
Keywords: autophagy; degradation; proteasome; ubiquitin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
How the proteasome is degraded.Proc Natl Acad Sci U S A. 2016 Nov 22;113(47):13266-13268. doi: 10.1073/pnas.1616535113. Epub 2016 Nov 11. Proc Natl Acad Sci U S A. 2016. PMID: 27837026 Free PMC article. No abstract available.
-
p62 at the crossroad of the ubiquitin-proteasome system and autophagy.Oncotarget. 2016 Dec 20;7(51):83833-83834. doi: 10.18632/oncotarget.13805. Oncotarget. 2016. PMID: 27974671 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
