

Quantifying the Assembly of Multicomponent Molecular Machines by Single-Molecule Total Internal Reflection Fluorescence Microscopy
- PMID: 27793278
- PMCID: PMC5403009
- DOI: 10.1016/bs.mie.2016.08.019
Quantifying the Assembly of Multicomponent Molecular Machines by Single-Molecule Total Internal Reflection Fluorescence Microscopy
Abstract
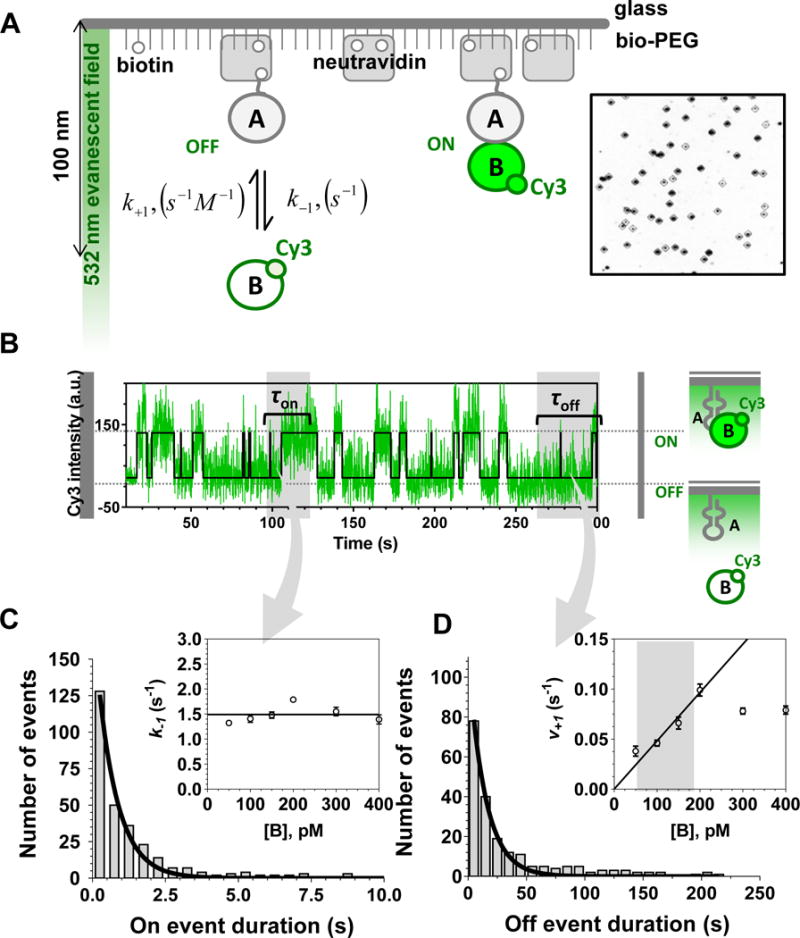
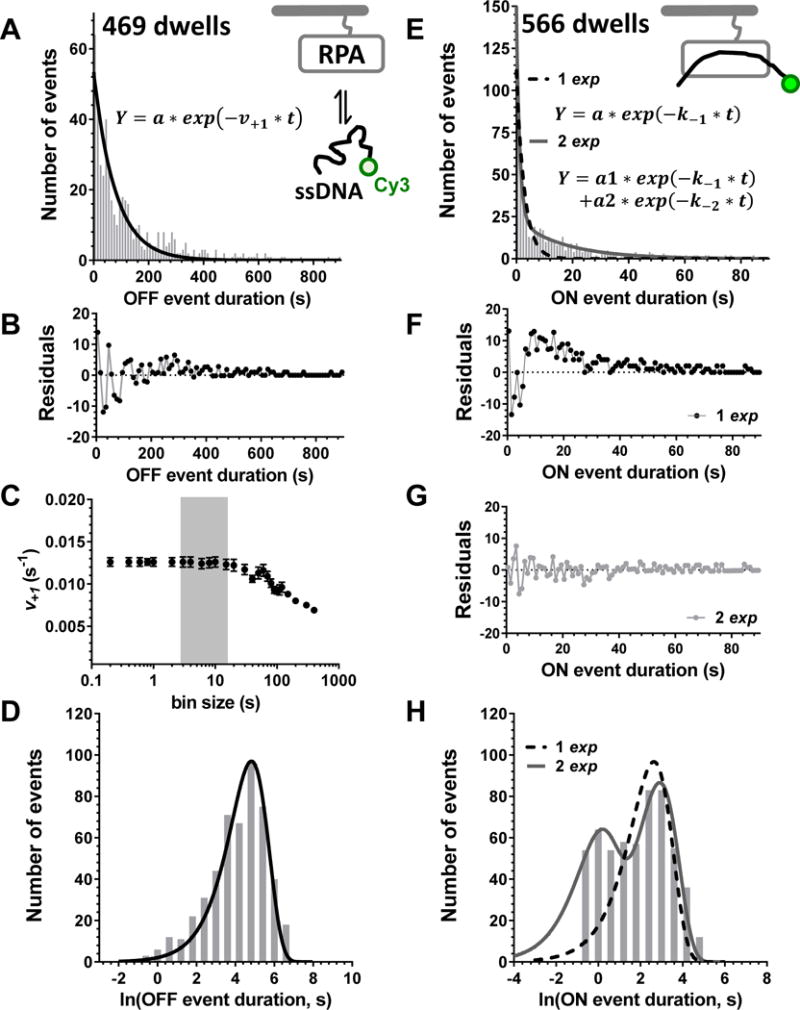
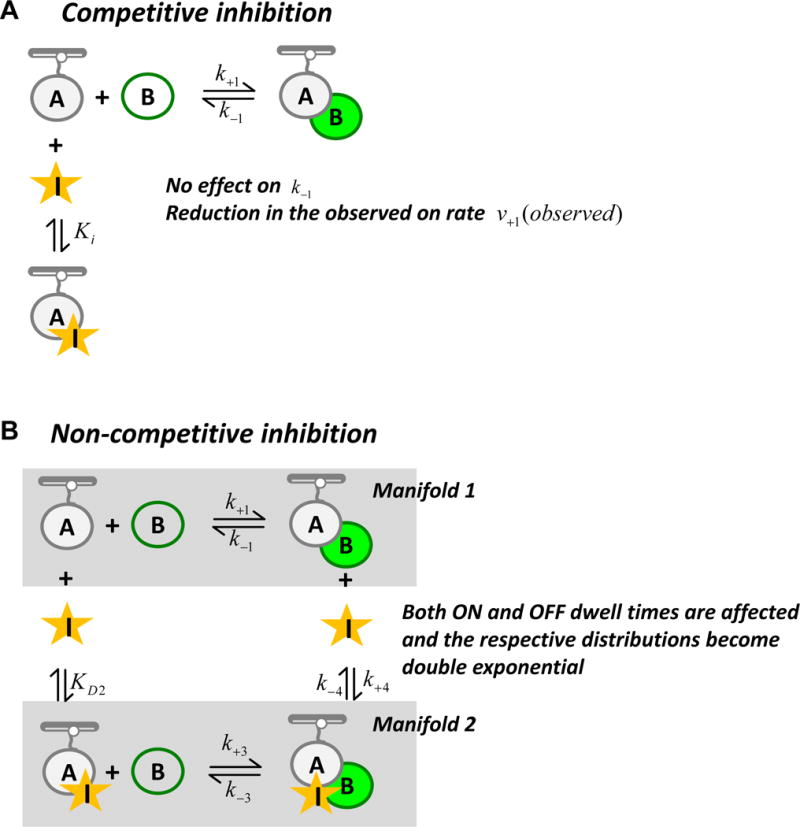
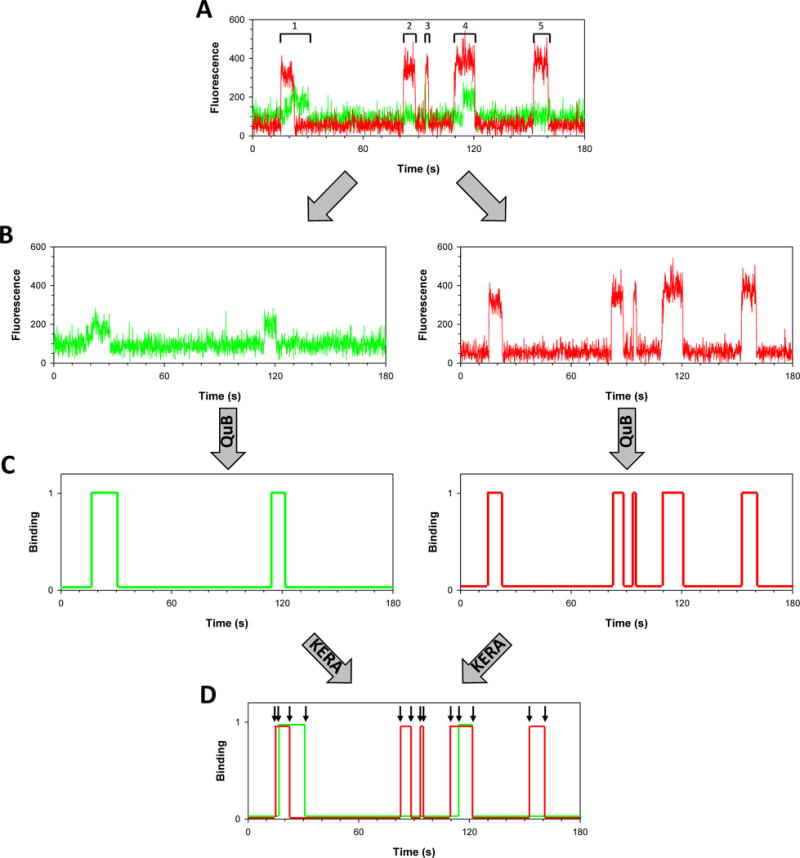
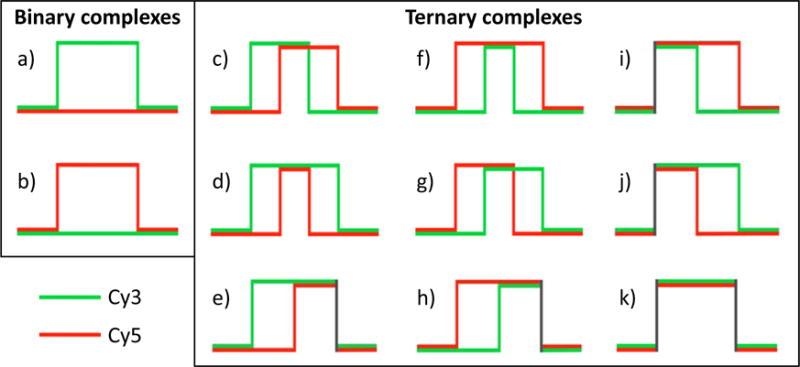
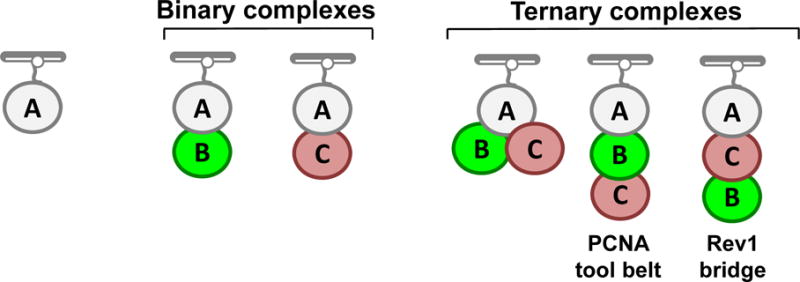
Large, dynamic macromolecular complexes play essential roles in many cellular processes. Knowing how the components of these complexes associate with one another and undergo structural rearrangements is critical to understanding how they function. Single-molecule total internal reflection fluorescence (TIRF) microscopy is a powerful approach for addressing these fundamental issues. In this article, we first discuss single-molecule TIRF microscopes and strategies to immobilize and fluorescently label macromolecules. We then review the use of single-molecule TIRF microscopy to study the formation of binary macromolecular complexes using one-color imaging and inhibitors. We conclude with a discussion of the use of TIRF microscopy to examine the formation of higher-order (i.e., ternary) complexes using multicolor setups. The focus throughout this article is on experimental design, controls, data acquisition, and data analysis. We hope that single-molecule TIRF microscopy, which has largely been the province of specialists, will soon become as common in the tool box of biophysicists and biochemists as structural approaches have become today.
Keywords: Colocalization single-molecule imaging; Competitive and noncompetitive inhibition; Kinetic event resolving algorithm; Protein–protein and protein–nucleic acid interaction; Single-molecule fluorescence; Total internal reflection fluorescence microscopy.
© 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- AGGARWAL V, HA T. Single-molecule pull-down (SiMPull) for new-age biochemistry: methodology and biochemical applications of single-molecule pull-down (SiMPull) for probing biomolecular interactions in crude cell extracts. Bioessays. 2014;36:1109–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
