

Strigolactone regulates shoot development through a core signalling pathway
- PMID: 27793831
- PMCID: PMC5200909
- DOI: 10.1242/bio.021402
Strigolactone regulates shoot development through a core signalling pathway
Abstract
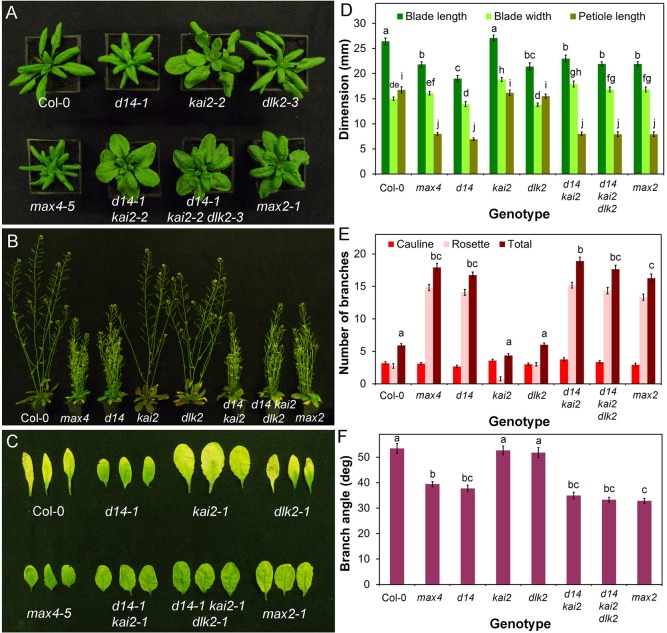
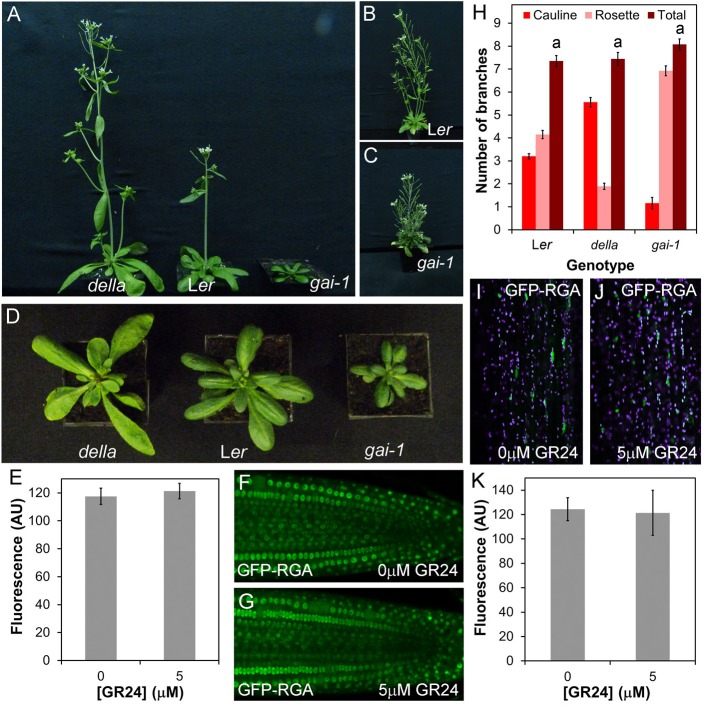
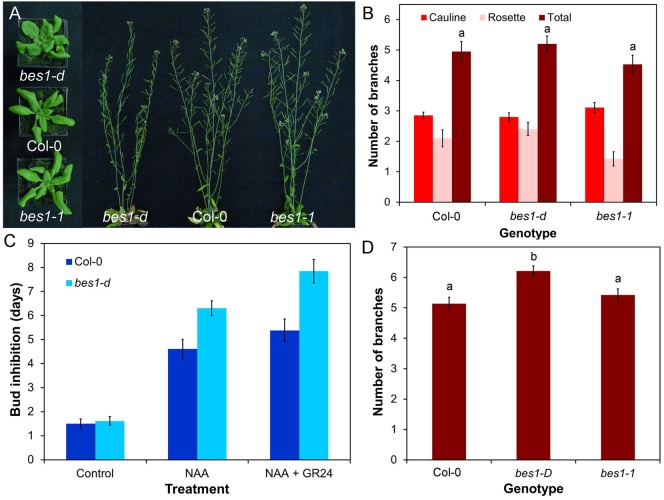
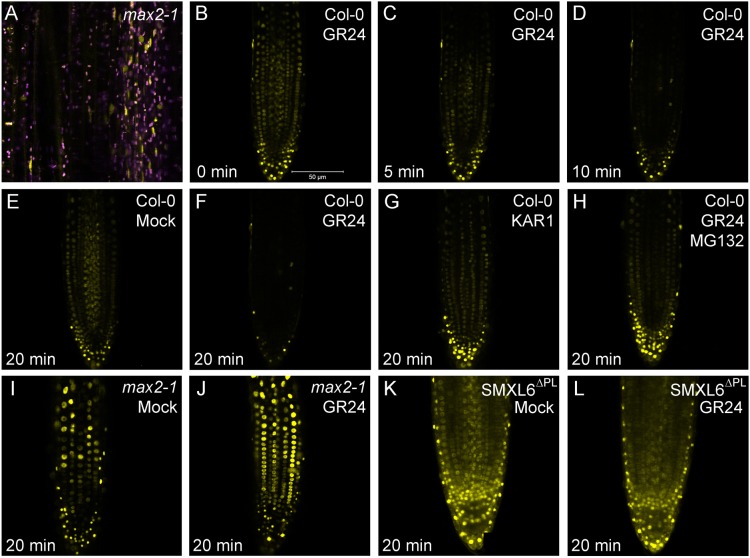
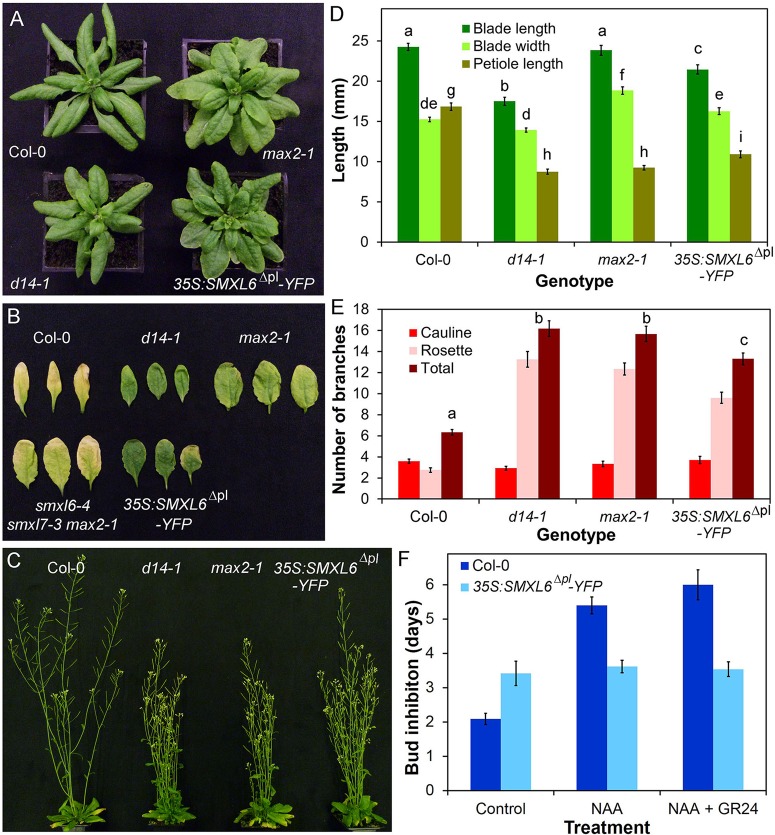
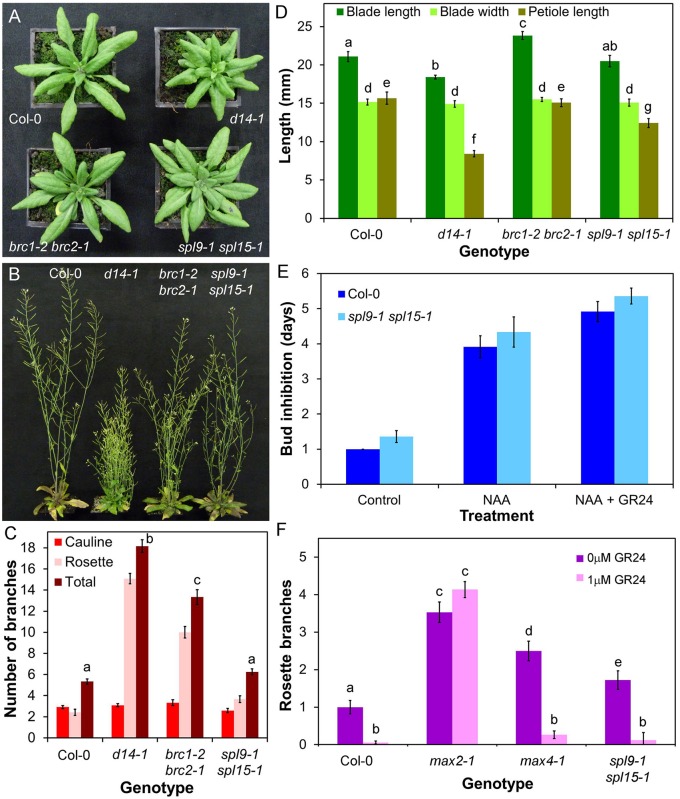
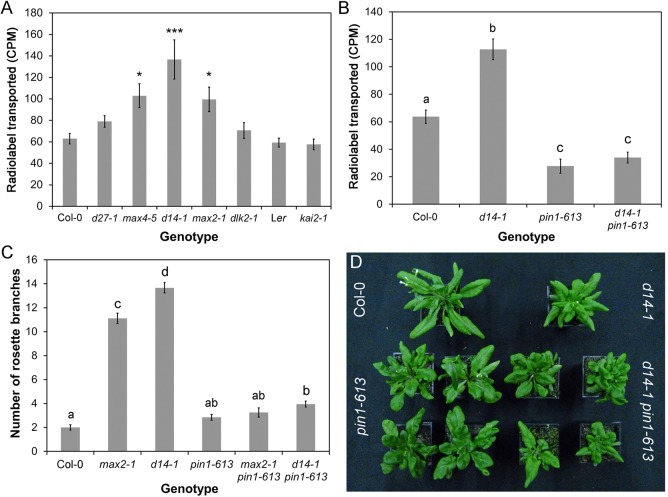
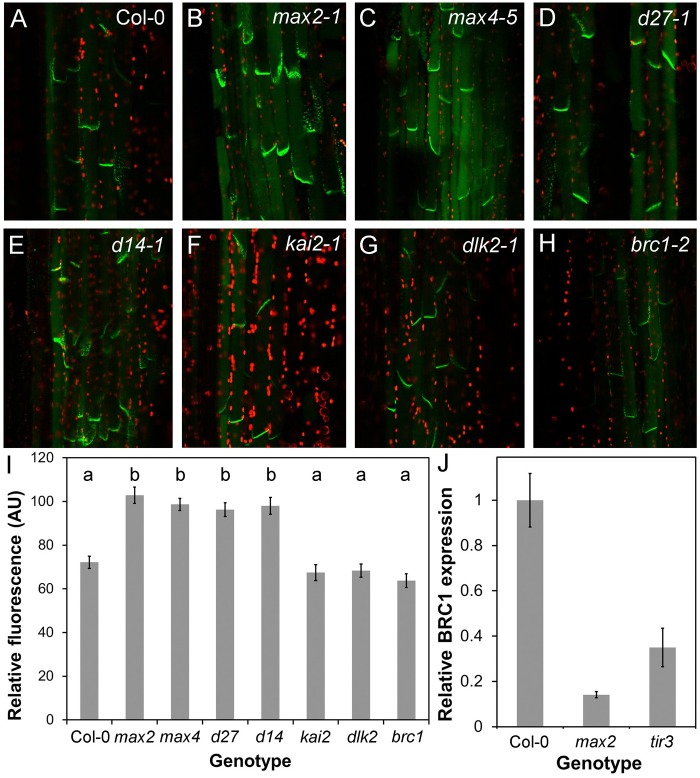
Strigolactones are a recently identified class of hormone that regulate multiple aspects of plant development. The DWARF14 (D14) α/β fold protein has been identified as a strigolactone receptor, which can act through the SCFMAX2 ubiquitin ligase, but the universality of this mechanism is not clear. Multiple proteins have been suggested as targets for strigolactone signalling, including both direct proteolytic targets of SCFMAX2, and downstream targets. However, the relevance and importance of these proteins to strigolactone signalling in many cases has not been fully established. Here we assess the contribution of these targets to strigolactone signalling in adult shoot developmental responses. We find that all examined strigolactone responses are regulated by SCFMAX2 and D14, and not by other D14-like proteins. We further show that all examined strigolactone responses likely depend on degradation of SMXL proteins in the SMXL6 clade, and not on the other proposed proteolytic targets BES1 or DELLAs. Taken together, our results suggest that in the adult shoot, the dominant mode of strigolactone signalling is D14-initiated, MAX2-mediated degradation of SMXL6-related proteins. We confirm that the BRANCHED1 transcription factor and the PIN-FORMED1 auxin efflux carrier are plausible downstream targets of this pathway in the regulation of shoot branching, and show that BRC1 likely acts in parallel to PIN1.
Keywords: Shoot branching; Signal transduction; Strigolactone.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
-
- Abe S., Sado A., Tanaka K., Kisugi T., Asami K., Ota S., Kim H. I., Yoneyama K., Xie X., Ohnishi T. et al. (2014). Carlactone is converted to carlactonoic acid by MAX1 in Arabidopsis and its methyl ester can directly interact with AtD14 in vitro. Proc. Natl. Acad. Sci. USA 111, 18084-18089. 10.1073/pnas.1410801111 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
