

SIRT6 delays cellular senescence by promoting p27Kip1 ubiquitin-proteasome degradation
- PMID: 27794562
- PMCID: PMC5115890
- DOI: 10.18632/aging.101038
SIRT6 delays cellular senescence by promoting p27Kip1 ubiquitin-proteasome degradation
Abstract
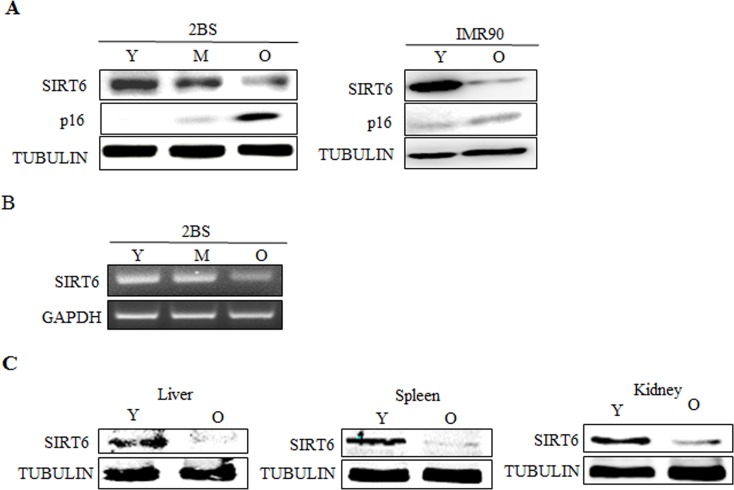
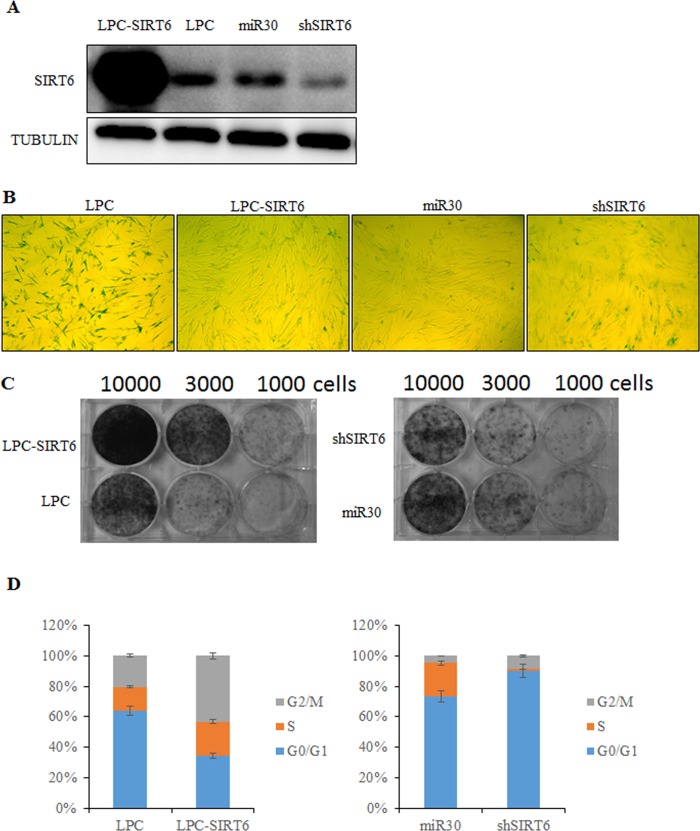
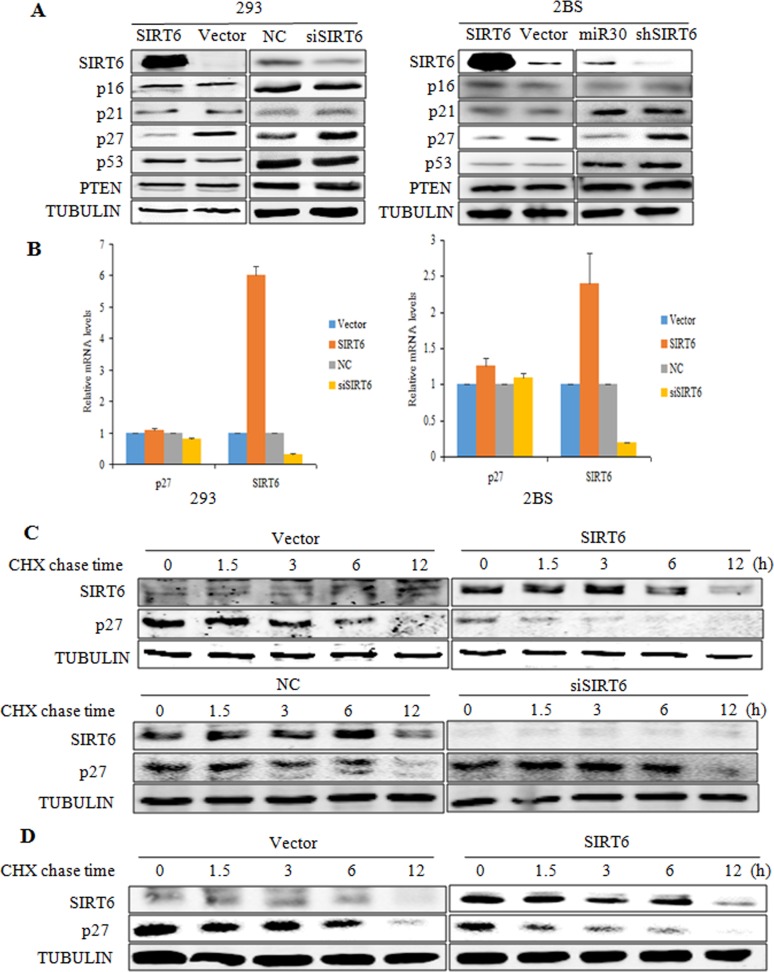
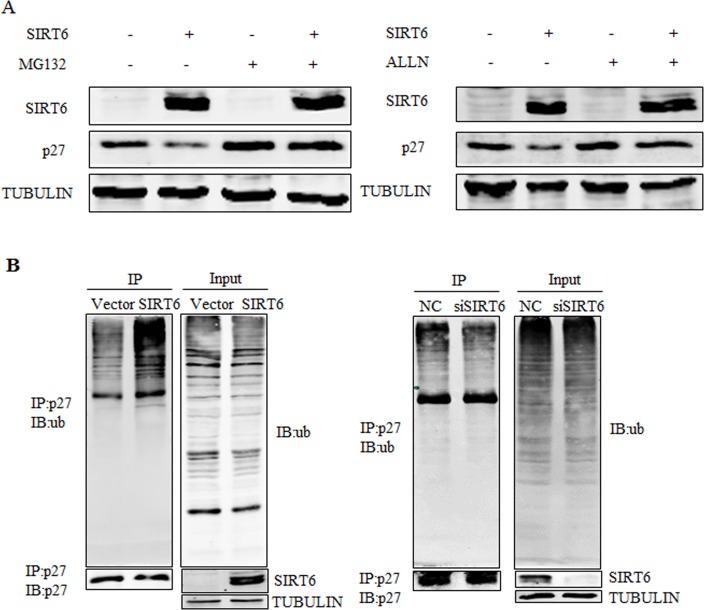
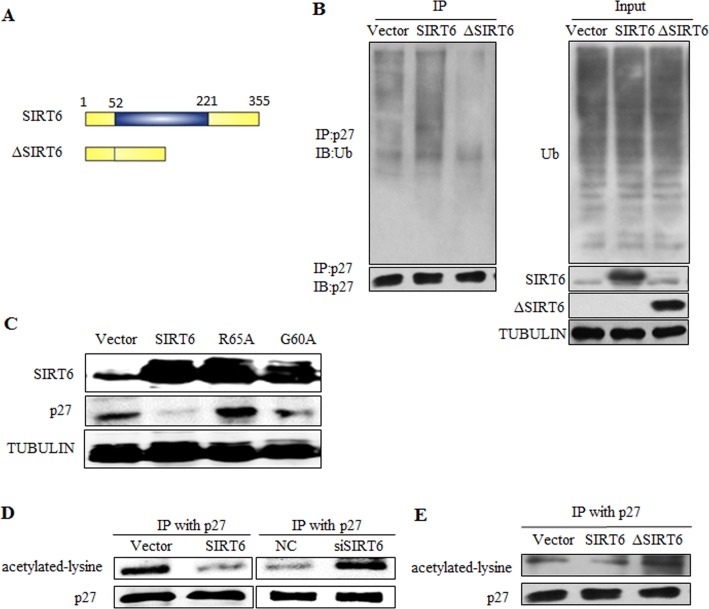
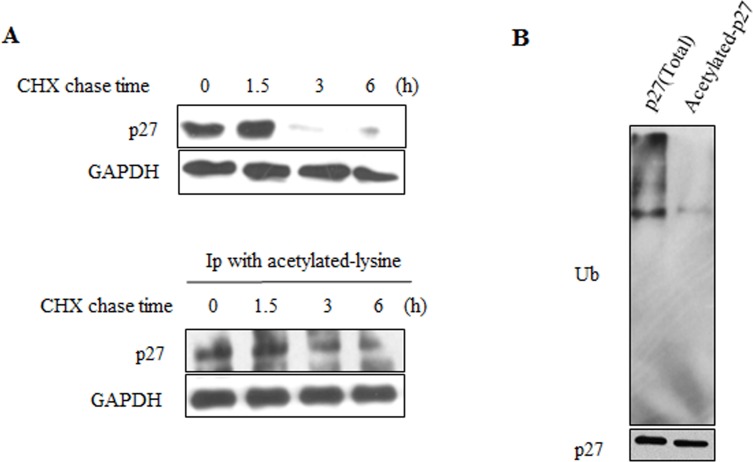
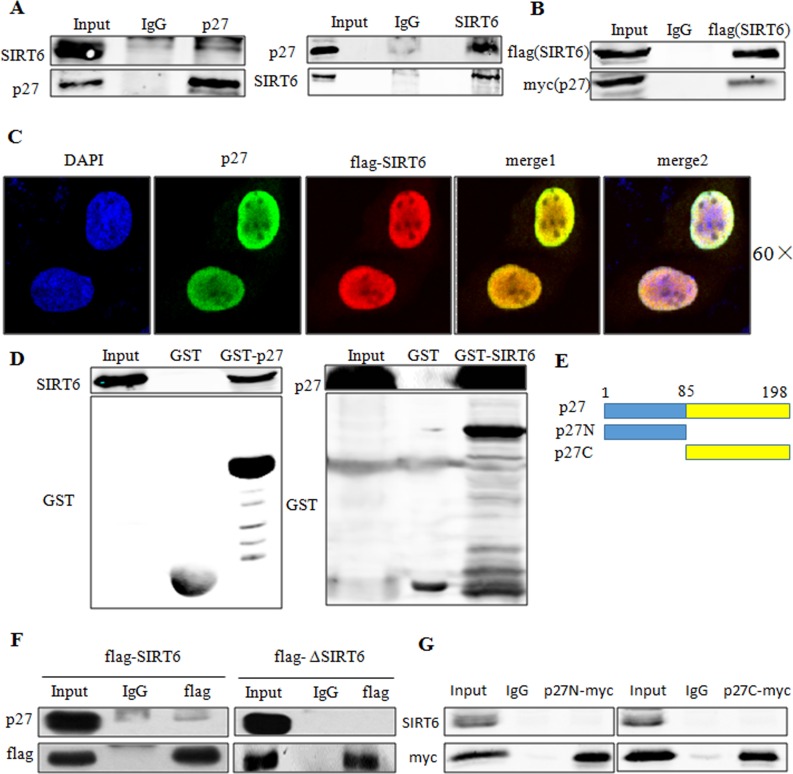
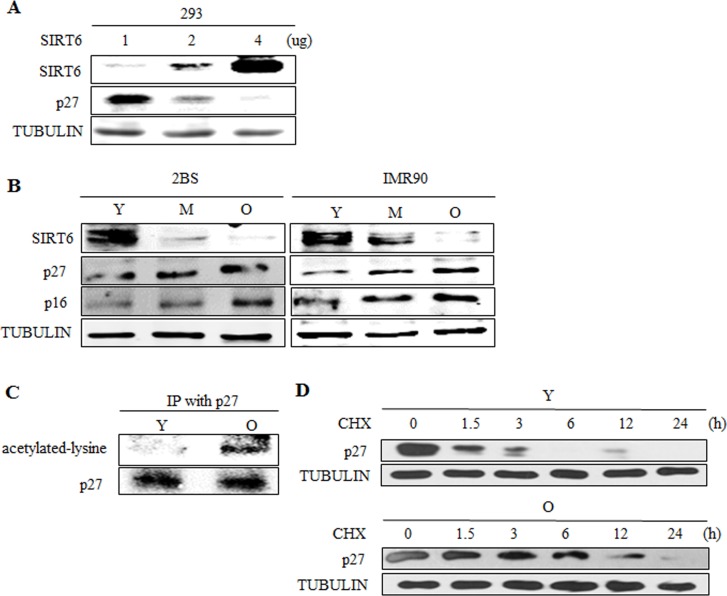
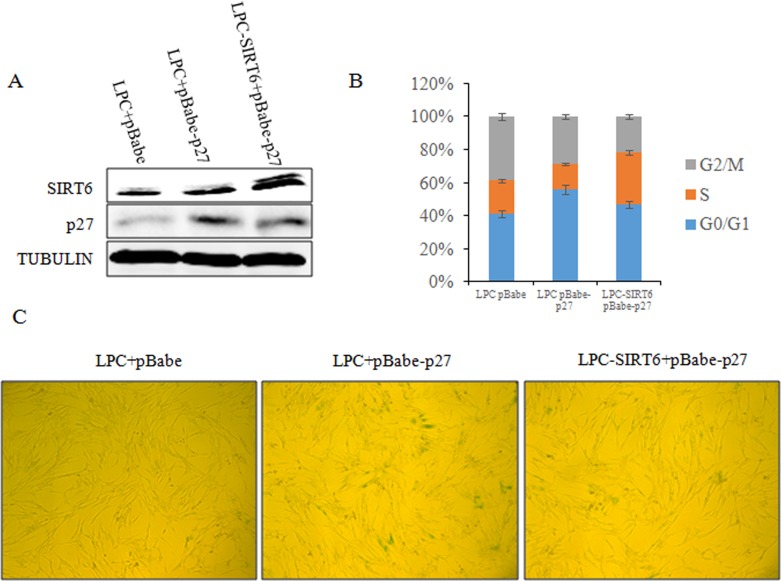
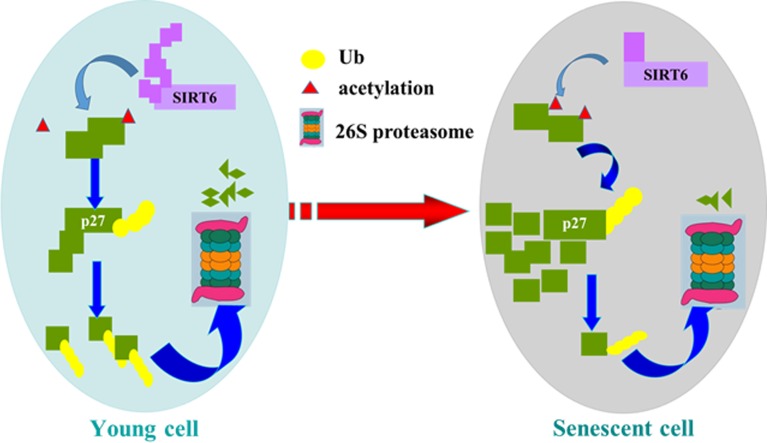
Sirtuin6(SIRT6) has been implicated as a key factor in aging and aging-related diseases. However, the role of SIRT6 in cellular senescence has not been fully understood. Here, we show that SIRT6 repressed the expression of p27Kip1 (p27) in cellular senescence. The expression of SIRT6 was reduced during cellular senescence, whereas enforced SIRT6 expression promoted cell proliferation and antagonized cellular senescence. In addition, we demonstrated that SIRT6 promoted p27 degradation by proteasome and SIRT6 decreased the acetylation level and protein half-life of p27. p27 acetylation increased its protein stability. Furthermore, SIRT6 directly interacted with p27. Importantly, p27 was strongly acetylated and had a prolonged protein half-life with the reduction of SIRT6 when cells were senescent, compared with those young cells. Finally, SIRT6 markedly rescued senescence induced by p27. Our findings indicate that SIRT6 decreases p27 acetylation, leading to its degradation via ubiquitin-proteasome pathway and then delays cellular senescence.
Keywords: SIRT6; acetylation; cellular senescence; p27kip1; ubiquitination.
Conflict of interest statement
The authors have no conflict of interests to declare.
Figures
Similar articles
-
WW domain-containing E3 ubiquitin protein ligase 1 (WWP1) delays cellular senescence by promoting p27(Kip1) degradation in human diploid fibroblasts.J Biol Chem. 2011 Sep 23;286(38):33447-56. doi: 10.1074/jbc.M111.225565. Epub 2011 Jul 27. J Biol Chem. 2011. PMID: 21795702 Free PMC article.
-
Damage-specific DNA binding protein 1 (DDB1) is involved in ubiquitin-mediated proteolysis of p27Kip1 in response to UV irradiation.Biochimie. 2011 May;93(5):867-75. doi: 10.1016/j.biochi.2010.12.017. Epub 2011 Jan 13. Biochimie. 2011. PMID: 21237244
-
Interferon-γ-induced p27KIP1 binds to and targets MYC for proteasome-mediated degradation.Oncotarget. 2016 Jan 19;7(3):2837-54. doi: 10.18632/oncotarget.6693. Oncotarget. 2016. PMID: 26701207 Free PMC article.
-
p27(Kip1) signaling: Transcriptional and post-translational regulation.Int J Biochem Cell Biol. 2015 Nov;68:9-14. doi: 10.1016/j.biocel.2015.08.005. Epub 2015 Aug 14. Int J Biochem Cell Biol. 2015. PMID: 26279144 Review.
-
Post-translational modification of cellular proteins by ubiquitin and ubiquitin-like molecules: role in cellular senescence and aging.Adv Exp Med Biol. 2010;694:172-96. doi: 10.1007/978-1-4419-7002-2_13. Adv Exp Med Biol. 2010. PMID: 20886764 Review.
Cited by
-
Connecting epigenetics and inflammation in vascular senescence: state of the art, biomarkers and senotherapeutics.Front Genet. 2024 Feb 26;15:1345459. doi: 10.3389/fgene.2024.1345459. eCollection 2024. Front Genet. 2024. PMID: 38469117 Free PMC article. Review.
-
SIRT6, a Mammalian Deacylase with Multitasking Abilities.Physiol Rev. 2020 Jan 1;100(1):145-169. doi: 10.1152/physrev.00030.2018. Epub 2019 Aug 22. Physiol Rev. 2020. PMID: 31437090 Free PMC article. Review.
-
Sporoderm-Removed Ganoderma lucidum Spore Powder May Suppress the Proliferation, Migration, and Invasion of Esophageal Squamous Cell Carcinoma Cells Through PI3K/AKT/mTOR and Erk Pathway.Integr Cancer Ther. 2021 Jan-Dec;20:15347354211062157. doi: 10.1177/15347354211062157. Integr Cancer Ther. 2021. PMID: 34841952 Free PMC article.
-
SIRT6-PAI-1 axis is a promising therapeutic target in aging-related bone metabolic disruption.Sci Rep. 2023 May 17;13(1):7991. doi: 10.1038/s41598-023-33297-7. Sci Rep. 2023. PMID: 37198221 Free PMC article.
-
Roles of Protein Post-Translational Modifications During Adipocyte Senescence.Int J Biol Sci. 2023 Oct 16;19(16):5245-5256. doi: 10.7150/ijbs.86404. eCollection 2023. Int J Biol Sci. 2023. PMID: 37928271 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
