

CLEC5A-Mediated Enhancement of the Inflammatory Response in Myeloid Cells Contributes to Influenza Virus Pathogenicity In Vivo
- PMID: 27795434
- PMCID: PMC5165214
- DOI: 10.1128/JVI.01813-16
CLEC5A-Mediated Enhancement of the Inflammatory Response in Myeloid Cells Contributes to Influenza Virus Pathogenicity In Vivo
Abstract
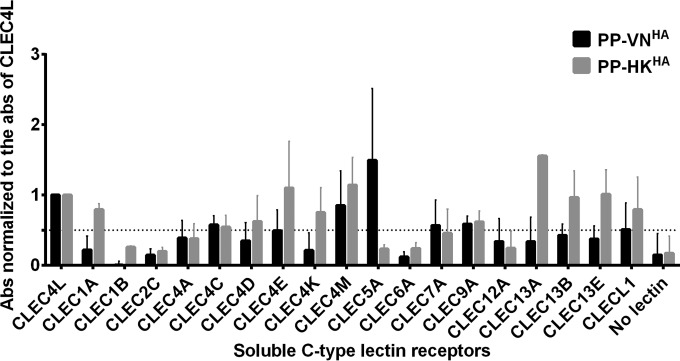
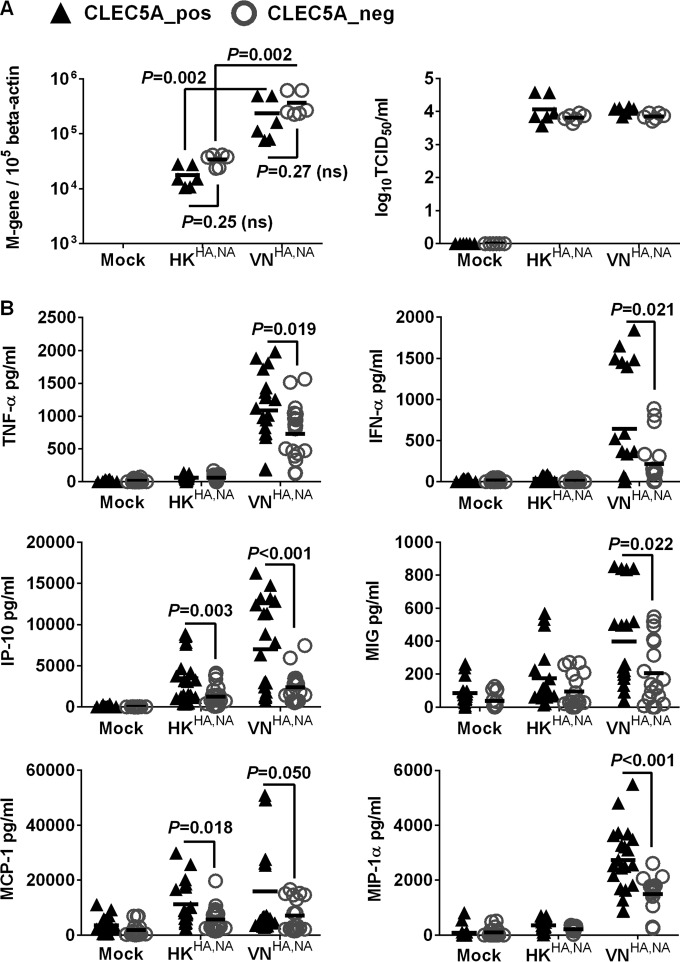
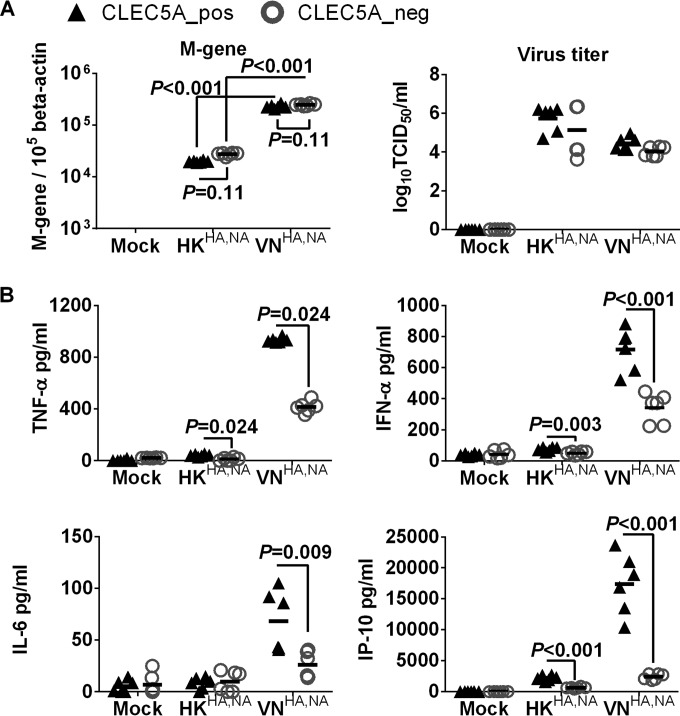
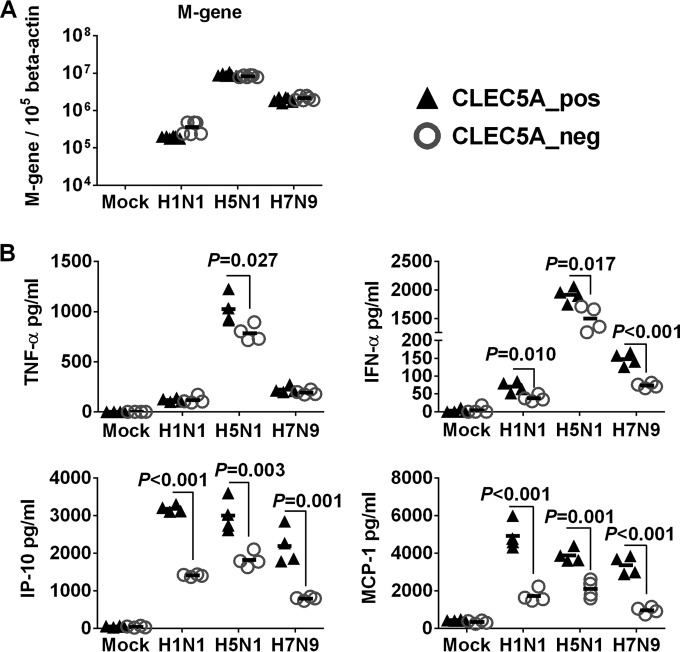
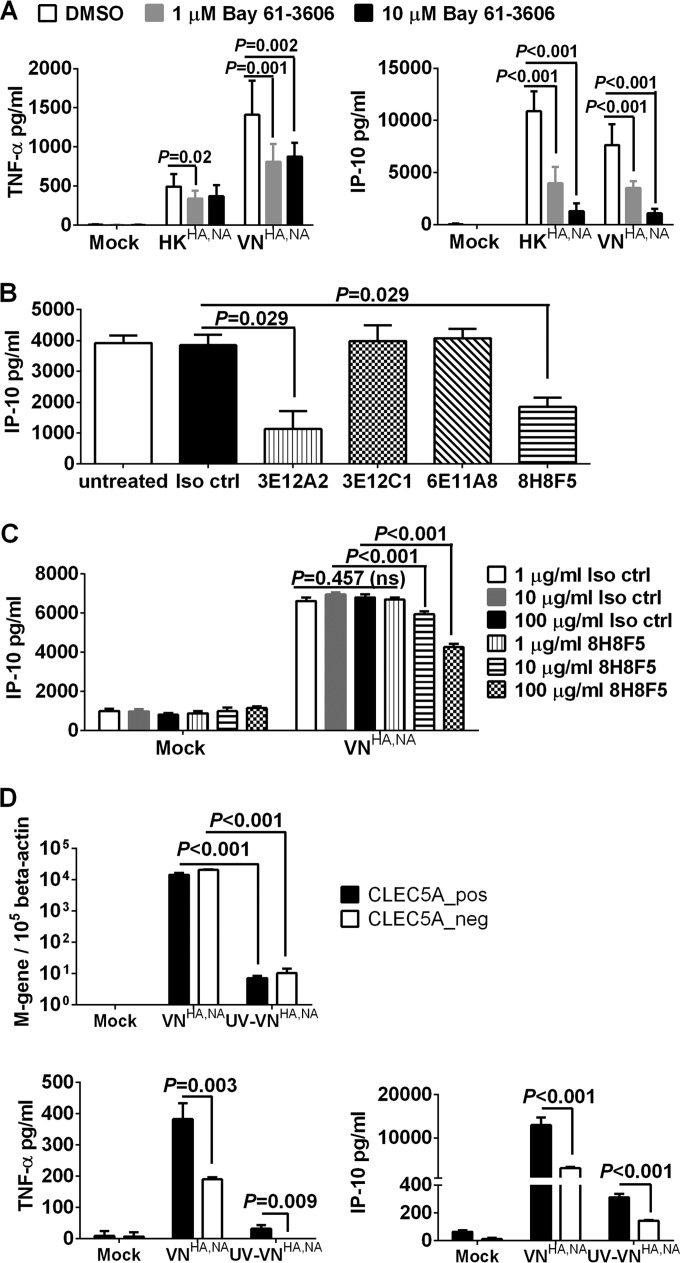
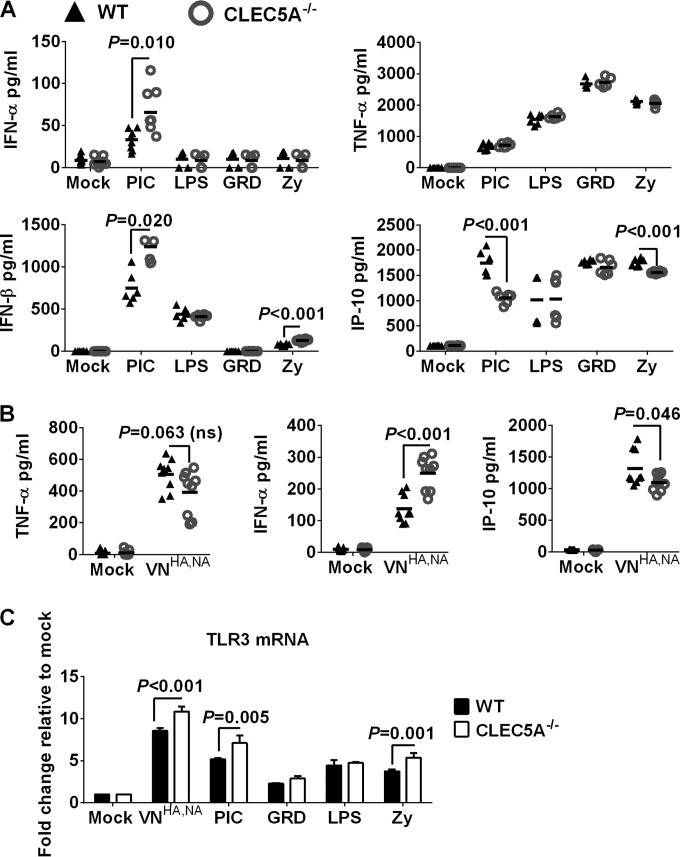
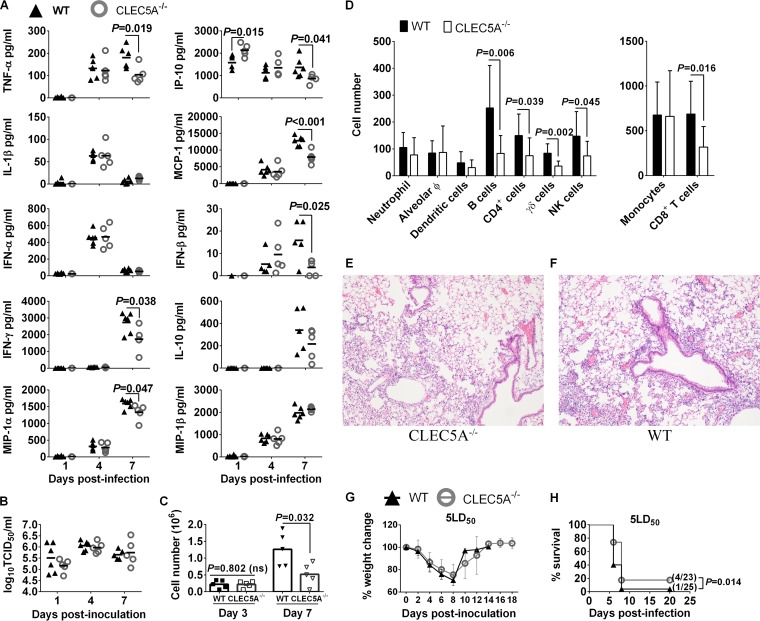
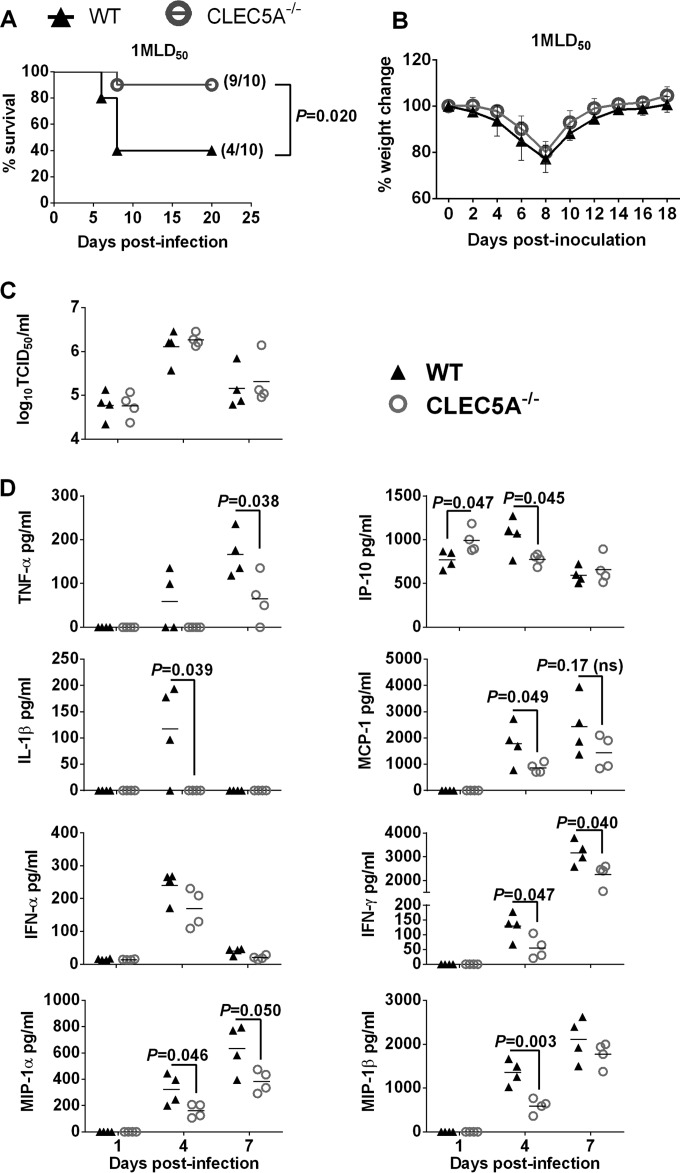
Human infections with influenza viruses exhibit mild to severe clinical outcomes as a result of complex virus-host interactions. Induction of inflammatory mediators via pattern recognition receptors may dictate subsequent host responses for pathogen clearance and tissue damage. We identified that human C-type lectin domain family 5 member A (CLEC5A) interacts with the hemagglutinin protein of influenza viruses expressed on lentiviral pseudoparticles through lectin screening. Silencing CLEC5A gene expression, blocking influenza-CLEC5A interactions with anti-CLEC5A antibodies, or dampening CLEC5A-mediated signaling using a spleen tyrosine kinase inhibitor consistently reduced the levels of proinflammatory cytokines produced by human macrophages without affecting the replication of influenza A viruses of different subtypes. Infection of bone marrow-derived macrophages from CLEC5A-deficient mice showed reduced levels of tumor necrosis factor alpha (TNF-α) and IP-10 but elevated alpha interferon (IFN-α) compared to those of wild-type mice. The heightened type I IFN response in the macrophages of CLEC5A-deficient mice was associated with upregulated TLR3 mRNA after treatment with double-stranded RNA. Upon lethal challenges with a recombinant H5N1 virus, CLEC5A-deficient mice showed reduced levels of proinflammatory cytokines, decreased immune cell infiltration in the lungs, and improved survival compared to the wild-type mice, despite comparable viral loads noted throughout the course of infection. The survival difference was more prominent at a lower dose of inoculum. Our results suggest that CLEC5A-mediated enhancement of the inflammatory response in myeloid cells contributes to influenza pathogenicity in vivo and may be considered a therapeutic target in combination with effective antivirals. Well-orchestrated host responses together with effective viral clearance are critical for optimal clinical outcome after influenza infections.
Importance: Multiple pattern recognition receptors work in synergy to sense viral RNA or proteins synthesized during influenza replication and mediate host responses for viral control. Well-orchestrated host responses may help to maintain the inflammatory response to minimize tissue damage while inducing an effective adaptive immune response for viral clearance. We identified that CLEC5A, a C-type lectin receptor which has previously been reported to mediate flavivirus-induced inflammatory responses, enhanced induction of proinflammatory cytokines and chemokines in myeloid cells after influenza infections. CLEC5A-deficient mice infected with influenza virus showed reduced inflammation in the lungs and improved survival compared to that of the wild-type mice despite comparable viral loads. The survival difference was more prominent at a lower dose of inoculum. Collectively, our results suggest that dampening CLEC5A-mediated inflammatory responses in myeloid cells reduces immunopathogenesis after influenza infections.
Keywords: C-type lectins; CLEC5A; influenza virus; macrophages; spleen tyrosine kinase (Syk).
Copyright © 2016 Teng et al.
Figures
References
-
- Everitt AR, Clare S, Pertel T, John SP, Wash RS, Smith SE, Chin CR, Feeley EM, Sims JS, Adams DJ, Wise HM, Kane L, Goulding D, Digard P, Anttila V, Baillie JK, Walsh TS, Hume DA, Palotie A, Xue Y, Colonna V, Tyler-Smith C, Dunning J, Gordon SB, GenISIS Investigators, MOSAIC Investigators, Smyth RL, Openshaw PJ, Dougan G, Brass AL, Kellam P. 2012. IFITM3 restricts the morbidity and mortality associated with influenza. Nature 484:519–523. doi: 10.1038/nature10921. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
