

Maintaining protein homeostasis: early and late endosomal dual recycling for the maintenance of intracellular pools of the plasma membrane protein Chs3
- PMID: 27798229
- PMCID: PMC5156543
- DOI: 10.1091/mbc.E16-04-0239
Maintaining protein homeostasis: early and late endosomal dual recycling for the maintenance of intracellular pools of the plasma membrane protein Chs3
Abstract
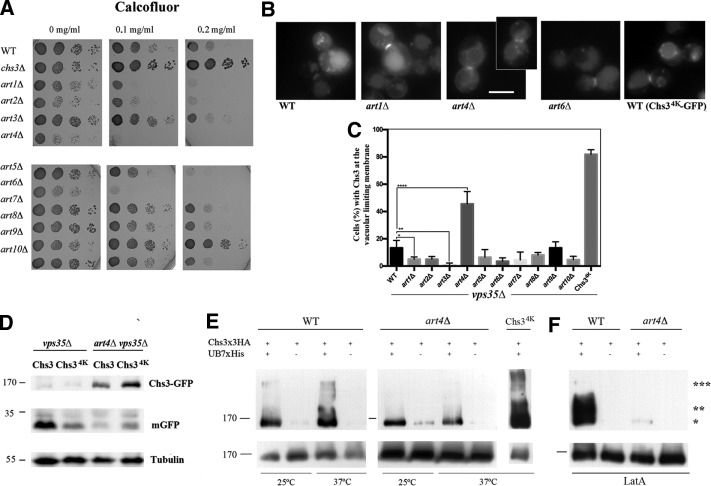
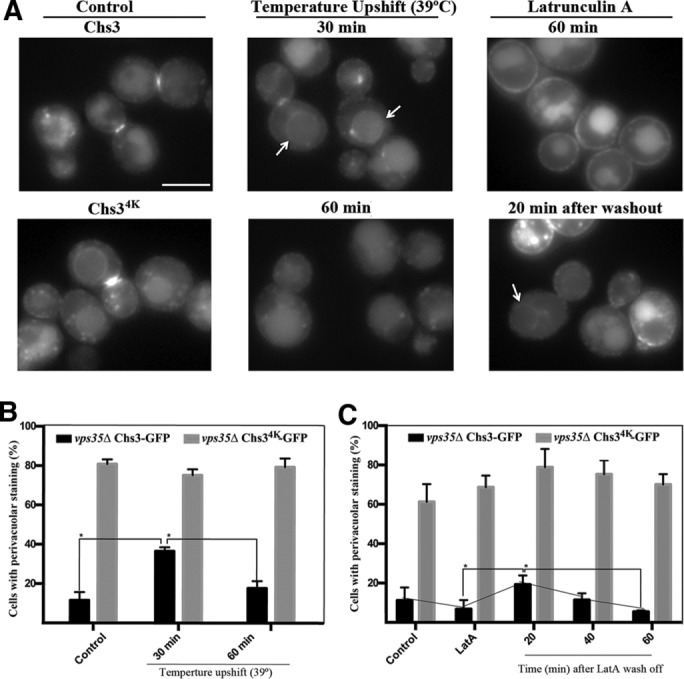
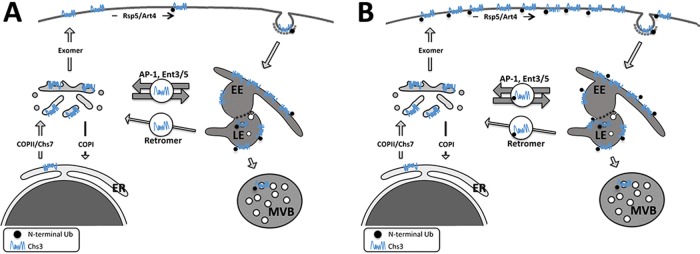
The major chitin synthase activity in yeast cells, Chs3, has become a paradigm in the study of the intracellular traffic of transmembrane proteins due to its tightly regulated trafficking. This includes an efficient mechanism for the maintenance of an extensive reservoir of Chs3 at the trans-Golgi network/EE, which allows for the timely delivery of the protein to the plasma membrane. Here we show that this intracellular reservoir of Chs3 is maintained not only by its efficient AP-1-mediated recycling, but also by recycling through the retromer complex, which interacts with Chs3 at a defined region in its N-terminal cytosolic domain. Moreover, the N-terminal ubiquitination of Chs3 at the plasma membrane by Rsp5/Art4 distinctly labels the protein and regulates its retromer-mediated recycling by enabling Chs3 to be recognized by the ESCRT machinery and degraded in the vacuole. Therefore the combined action of two independent but redundant endocytic recycling mechanisms, together with distinct labels for vacuolar degradation, determines the final fate of the intracellular traffic of the Chs3 protein, allowing yeast cells to regulate morphogenesis, depending on environmental constraints.
© 2016 Arcones et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
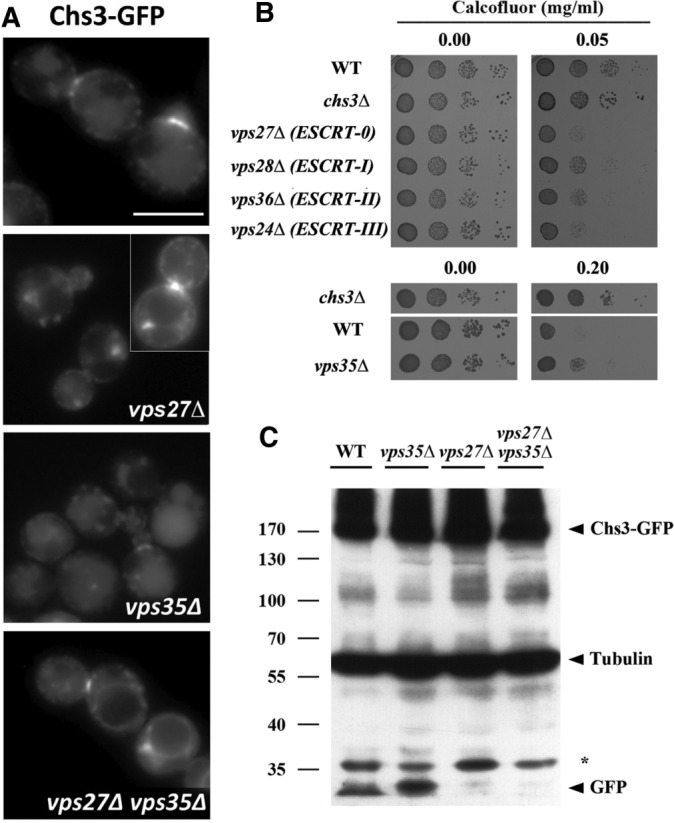
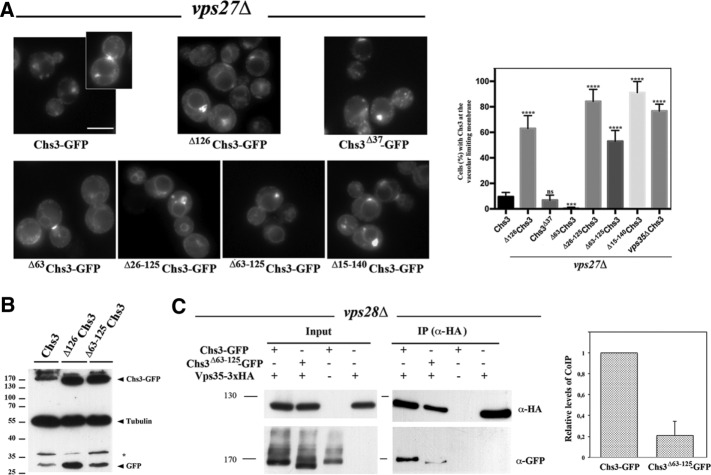
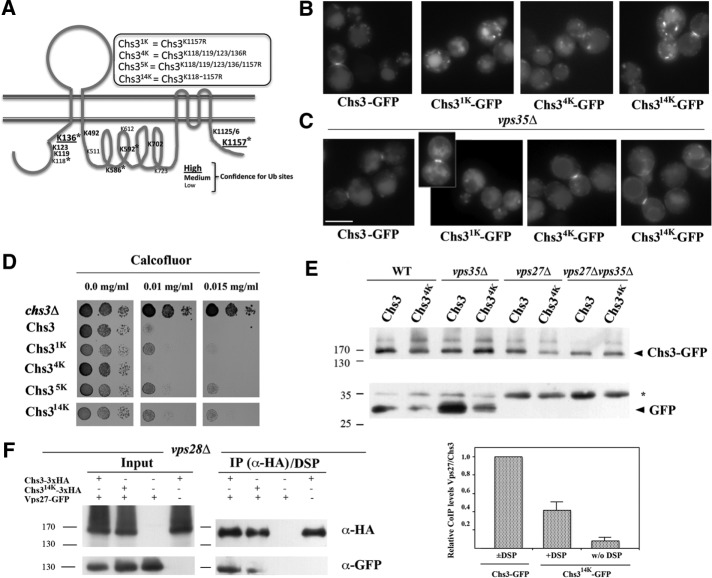
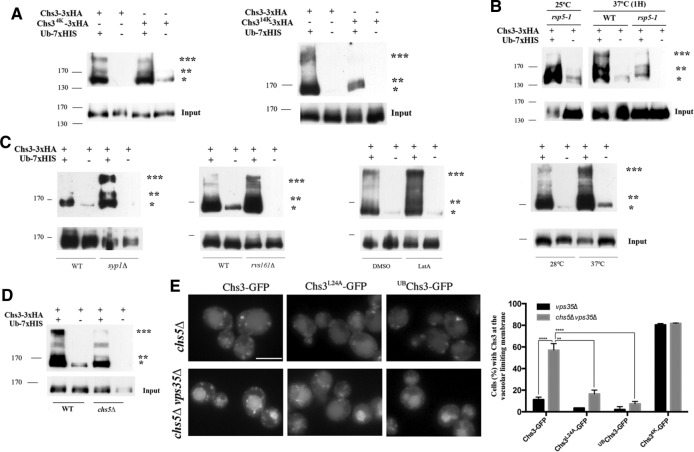
References
-
- Belgareh-Touzé N, Léon S, Erpapazoglou Z, Stawiecka-Mirota M, Urban-Grimal D, Haguenauer-Tsapis R. Versatile role of the yeast ubiquitin ligase Rsp5p in intracellular trafficking. Biochem Soc Trans. 2008;36:791–796. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
