

DUSP11 activity on triphosphorylated transcripts promotes Argonaute association with noncanonical viral microRNAs and regulates steady-state levels of cellular noncoding RNAs
- PMID: 27798849
- PMCID: PMC5066614
- DOI: 10.1101/gad.282616.116
DUSP11 activity on triphosphorylated transcripts promotes Argonaute association with noncanonical viral microRNAs and regulates steady-state levels of cellular noncoding RNAs
Abstract
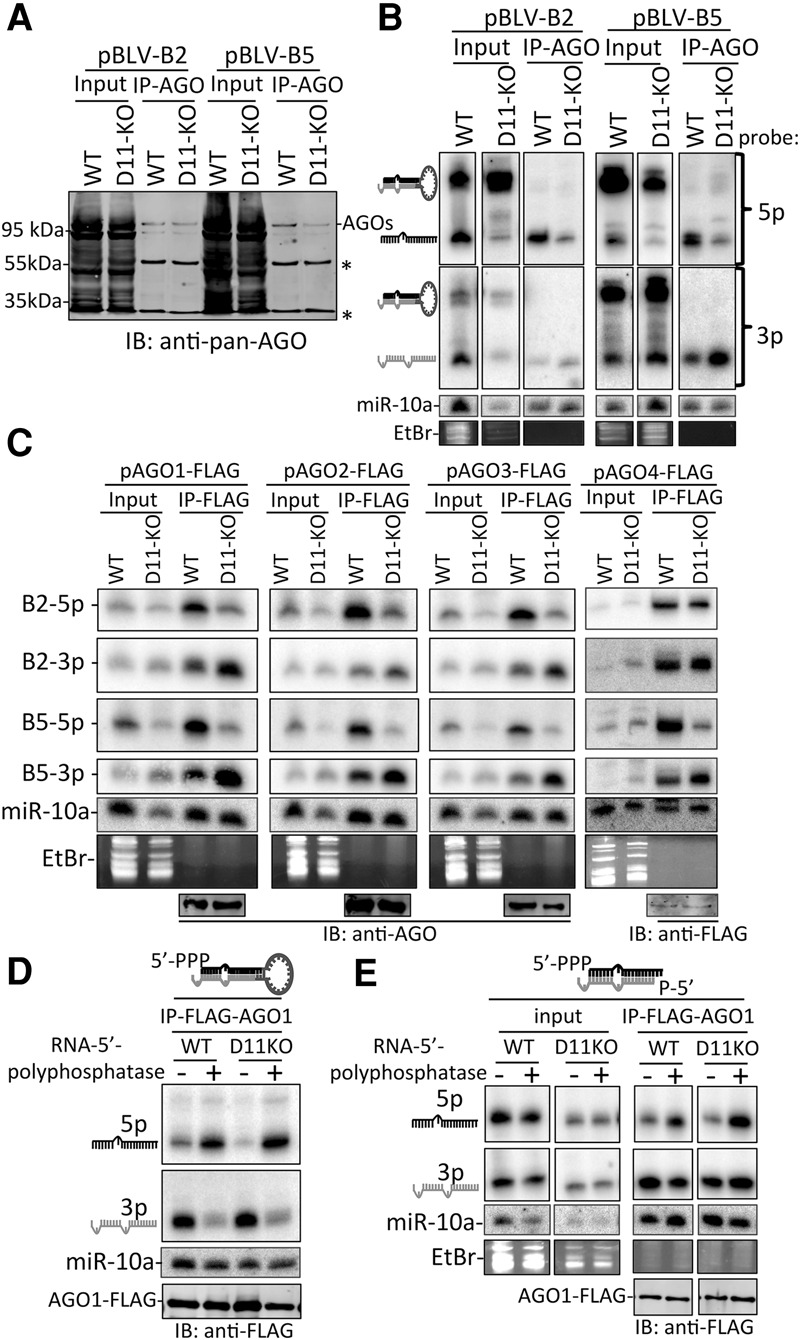
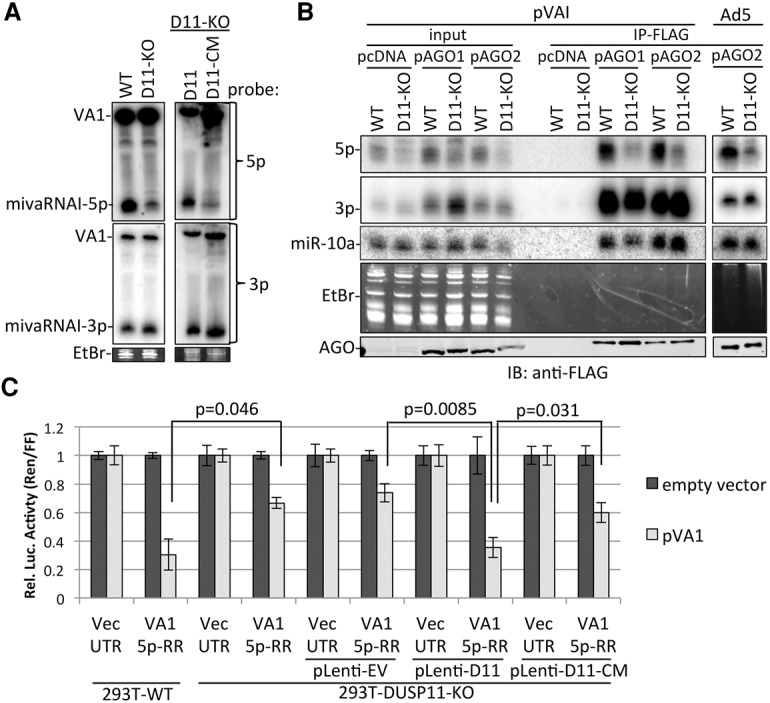
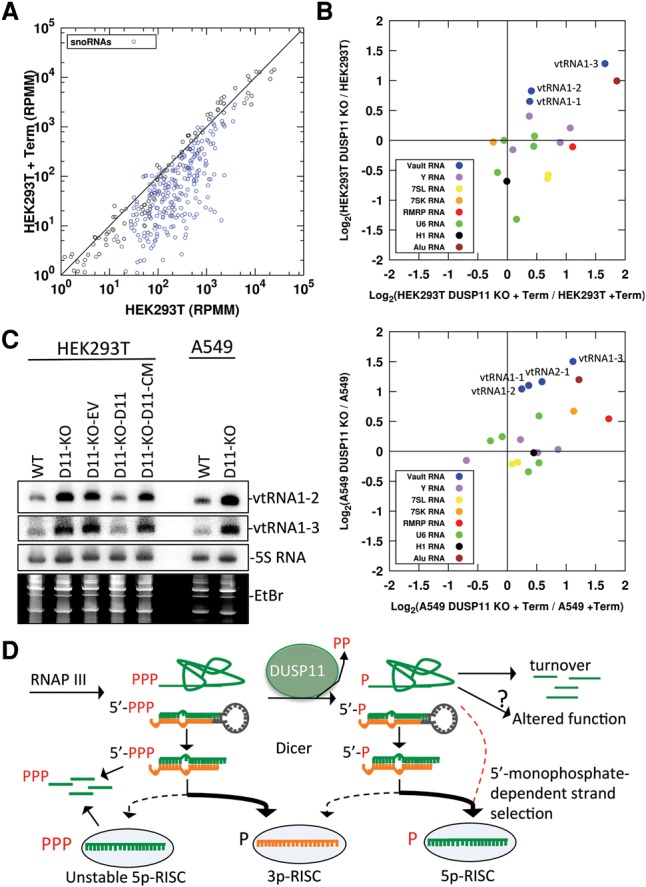
RNA silencing is a conserved eukaryotic gene expression regulatory mechanism mediated by small RNAs. In Caenorhabditis elegans, the accumulation of a distinct class of siRNAs synthesized by an RNA-dependent RNA polymerase (RdRP) requires the PIR-1 phosphatase. However, the function of PIR-1 in RNAi has remained unclear. Since mammals lack an analogous siRNA biogenesis pathway, an RNA silencing role for the mammalian PIR-1 homolog (dual specificity phosphatase 11 [DUSP11]) was unexpected. Here, we show that the RNA triphosphatase activity of DUSP11 promotes the RNA silencing activity of viral microRNAs (miRNAs) derived from RNA polymerase III (RNAP III) transcribed precursors. Our results demonstrate that DUSP11 converts the 5' triphosphate of miRNA precursors to a 5' monophosphate, promoting loading of derivative 5p miRNAs into Argonaute proteins via a Dicer-coupled 5' monophosphate-dependent strand selection mechanism. This mechanistic insight supports a likely shared function for PIR-1 in C. elegans Furthermore, we show that DUSP11 modulates the 5' end phosphate group and/or steady-state level of several host RNAP III transcripts, including vault RNAs and Alu transcripts. This study shows that steady-state levels of select noncoding RNAs are regulated by DUSP11 and defines a previously unknown portal for small RNA-mediated silencing in mammals, revealing that DUSP11-dependent RNA silencing activities are shared among diverse metazoans.
Keywords: Alu; BLV; DUSP11; RNAi; adenovirus; vault RNA.
© 2016 Burke et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
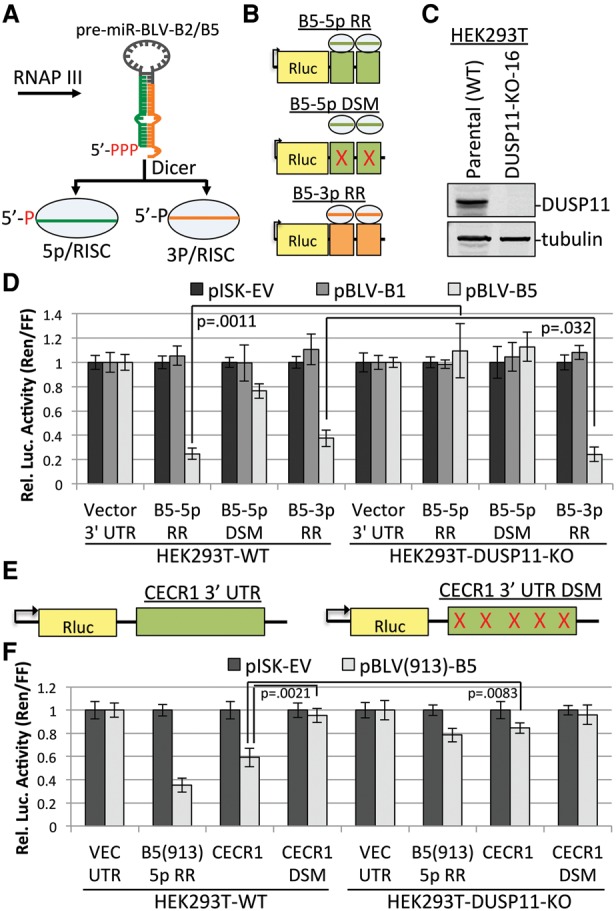
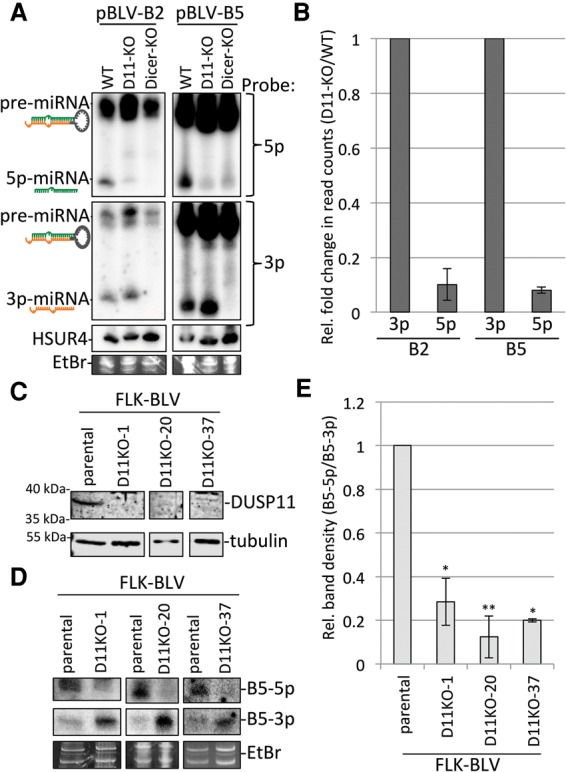
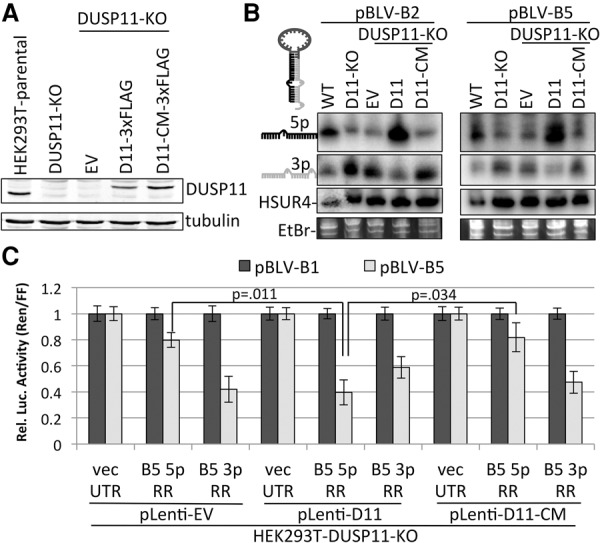
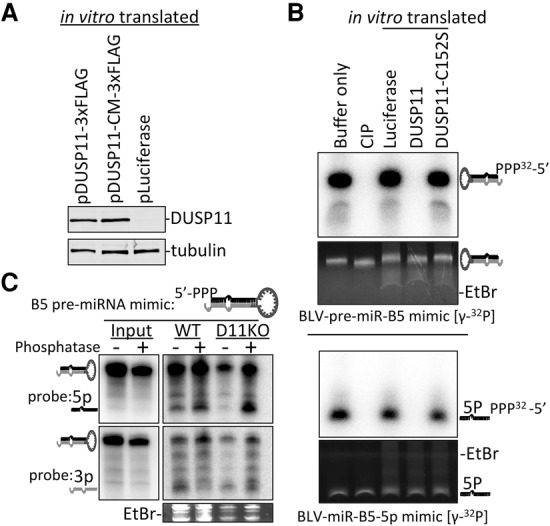
References
-
- Baek SE, Kim H, Kim KB, Yoon S, Choe J, Suh W, Jeong YJ, Cho YH, Kim DE. 2015. Dual effects of duplex RNA harboring 5′-terminal triphosphate on gene silencing and RIG-I mediated innate immune response. Biochem Biophys Res Commun 456: 591–597. - PubMed
-
- Bartel DP. 2004. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116: 281–297. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials