

HMGB2 orchestrates the chromatin landscape of senescence-associated secretory phenotype gene loci
- PMID: 27799366
- PMCID: PMC5100296
- DOI: 10.1083/jcb.201608026
HMGB2 orchestrates the chromatin landscape of senescence-associated secretory phenotype gene loci
Abstract
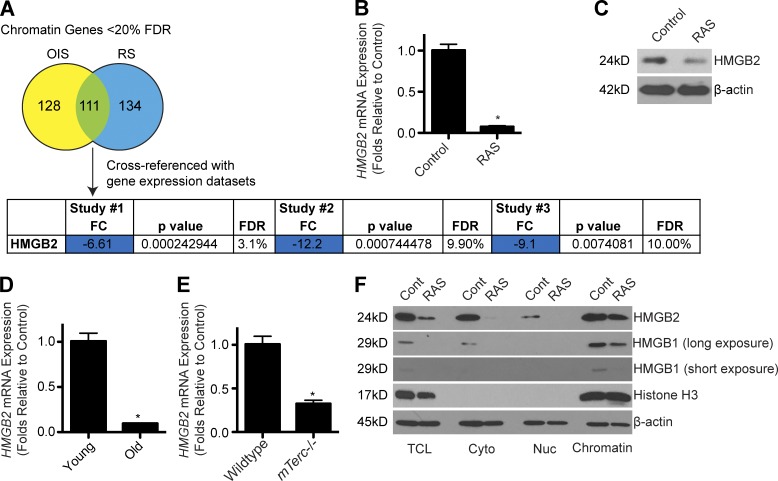
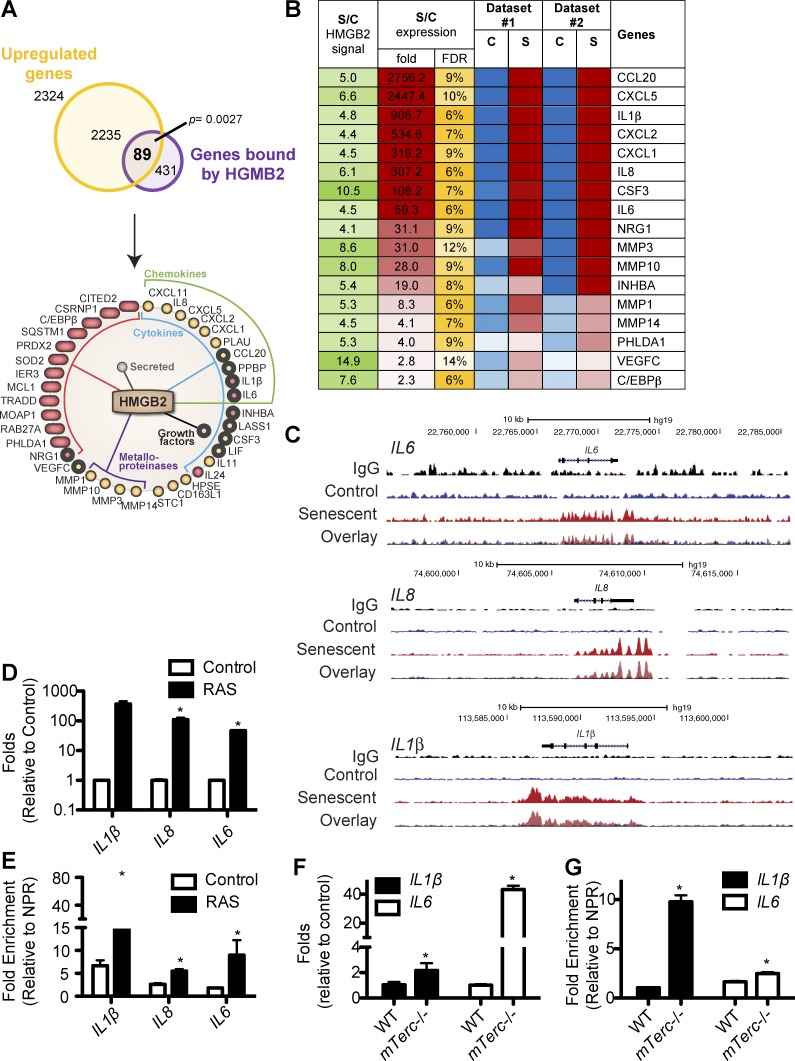
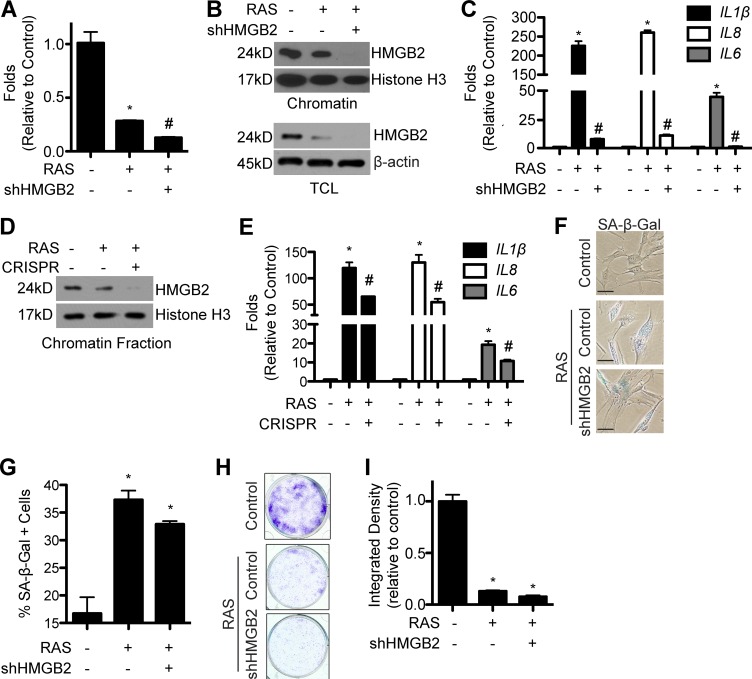
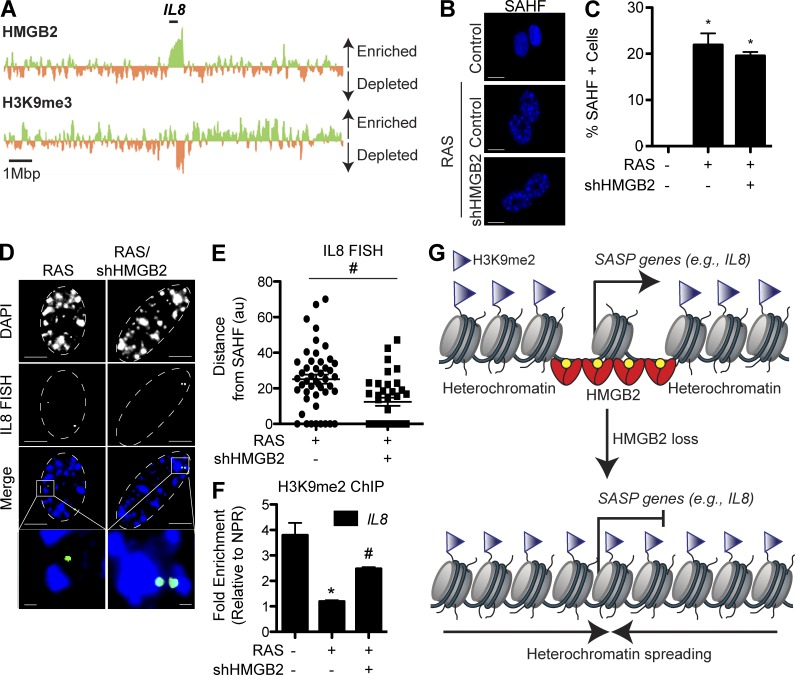
Cellular senescence is a stable cell growth arrest that is characterized by the silencing of proliferation-promoting genes through compaction of chromosomes into senescence-associated heterochromatin foci (SAHF). Paradoxically, senescence is also accompanied by increased transcription of certain genes encoding for secreted factors such as cytokines and chemokines, known as the senescence-associated secretory phenotype (SASP). How SASP genes are excluded from SAHF-mediated global gene silencing remains unclear. In this study, we report that high mobility group box 2 (HMGB2) orchestrates the chromatin landscape of SASP gene loci. HMGB2 preferentially localizes to SASP gene loci during senescence. Loss of HMGB2 during senescence blunts SASP gene expression by allowing for spreading of repressive heterochromatin into SASP gene loci. This correlates with incorporation of SASP gene loci into SAHF. Our results establish HMGB2 as a novel master regulator that orchestrates SASP through prevention of heterochromatin spreading to allow for exclusion of SASP gene loci from a global heterochromatin environment during senescence.
© 2016 Aird et al.
Figures
References
-
- Aird K.M., Zhang G., Li H., Tu Z., Bitler B.G., Garipov A., Wu H., Wei Z., Wagner S.N., Herlyn M., and Zhang R.. 2013. Suppression of nucleotide metabolism underlies the establishment and maintenance of oncogene-induced senescence. Cell Reports. 3:1252–1265. 10.1016/j.celrep.2013.03.004 - DOI - PMC - PubMed
-
- Biniossek M.L., Lechel A., Rudolph K.L., Martens U.M., and Zimmermann S.. 2013. Quantitative proteomic profiling of tumor cell response to telomere dysfunction using isotope-coded protein labeling (ICPL) reveals interaction network of candidate senescence markers. J. Proteomics. 91:515–535. 10.1016/j.jprot.2013.08.007 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
