

Junctophilin-2 in the nanoscale organisation and functional signalling of ryanodine receptor clusters in cardiomyocytes
- PMID: 27802169
- PMCID: PMC5201013
- DOI: 10.1242/jcs.196873
Junctophilin-2 in the nanoscale organisation and functional signalling of ryanodine receptor clusters in cardiomyocytes
Abstract
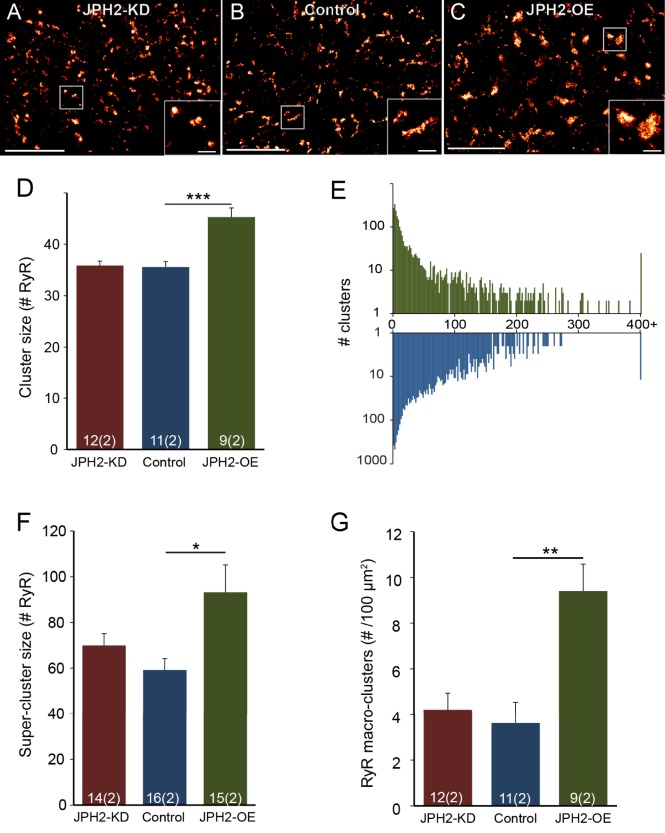
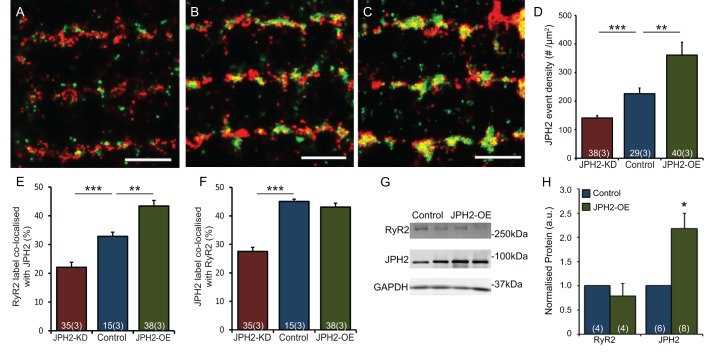
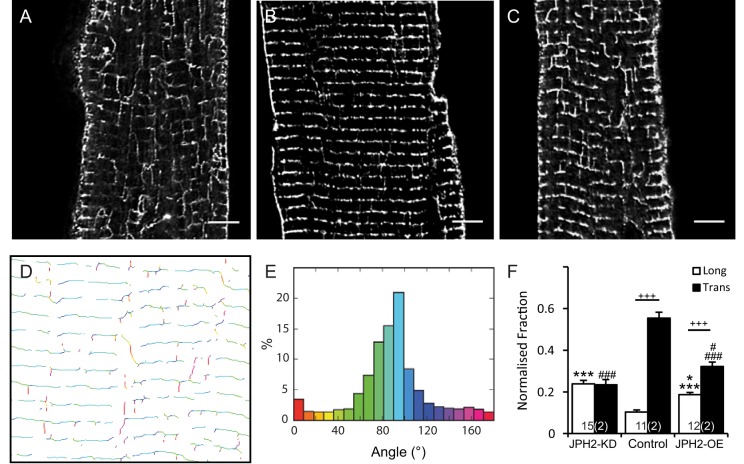
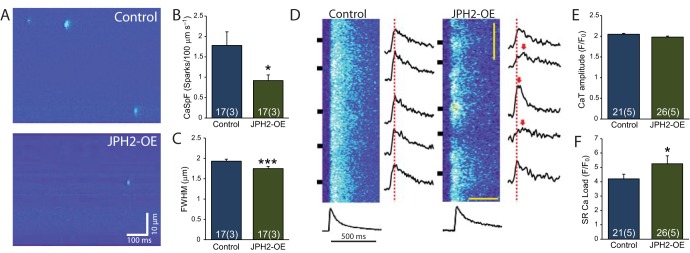
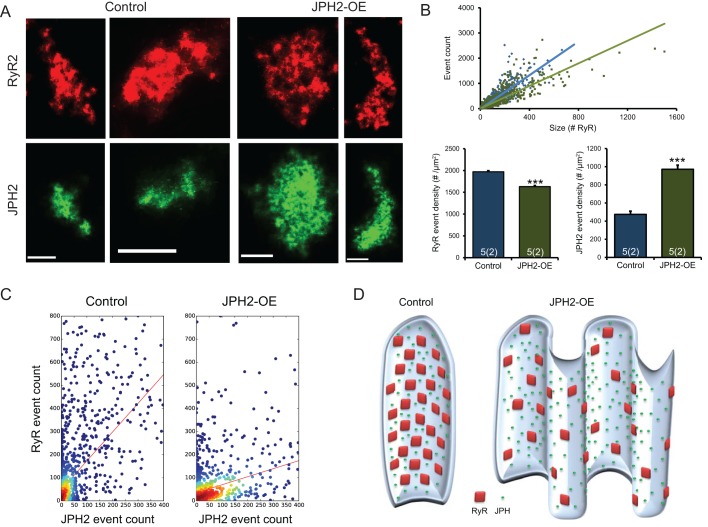
Signalling nanodomains requiring close contact between the plasma membrane and internal compartments, known as 'junctions', are fast communication hubs within excitable cells such as neurones and muscle. Here, we have examined two transgenic murine models probing the role of junctophilin-2, a membrane-tethering protein crucial for the formation and molecular organisation of sub-microscopic junctions in ventricular muscle cells of the heart. Quantitative single-molecule localisation microscopy showed that junctions in animals producing above-normal levels of junctophilin-2 were enlarged, allowing the re-organisation of the primary functional protein within it, the ryanodine receptor (RyR; in this paper, we use RyR to refer to the myocardial isoform RyR2). Although this change was associated with much enlarged RyR clusters that, due to their size, should be more excitable, functionally it caused a mild inhibition in the Ca2+ signalling output of the junctions (Ca2+ sparks). Analysis of the single-molecule densities of both RyR and junctophilin-2 revealed an ∼3-fold increase in the junctophilin-2 to RyR ratio. This molecular rearrangement is compatible with direct inhibition of RyR opening by junctophilin-2 to intrinsically stabilise the Ca2+ signalling properties of the junction and thus the contractile function of the cell.
Keywords: Ca2+ signalling; Excitation-contraction coupling; Junctophilin-2; Ryanodine receptor; Super-resolution imaging; dSTORM.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
X.H.T.W. is a founder and equity holder in Elex Biotech Llc.
Figures
References
-
- Baddeley D., Crossman D., Rossberger S., Cheyne J. E., Montgomery J. M., Jayasinghe I. D., Cremer C., Cannell M. B. and Soeller C. (2011). 4D super-resolution microscopy with conventional fluorophores and single wavelength excitation in optically thick cells and tissues. PLoS ONE 6, e20645 10.1371/journal.pone.0020645 - DOI - PMC - PubMed
-
- Beavers D. L., Wang W., Ather S., Voigt N., Garbino A., Dixit S. S., Landstrom A. P., Li N., Wang Q., Olivotto I. et al. (2013). Mutation E169K in Junctophilin-2 Causes Atrial Fibrillation Due to Impaired RyR2 Stabilization. J. Am. Coll. Cardiol. 62, 2010-2019. 10.1016/j.jacc.2013.06.052 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
