

Molecular Basis for Cohesin Acetylation by Establishment of Sister Chromatid Cohesion N-Acetyltransferase ESCO1
- PMID: 27803161
- PMCID: PMC5159507
- DOI: 10.1074/jbc.M116.752220
Molecular Basis for Cohesin Acetylation by Establishment of Sister Chromatid Cohesion N-Acetyltransferase ESCO1
Abstract
Protein acetylation is a prevalent posttranslational modification that is regulated by diverse acetyltransferase enzymes. Although histone acetyltransferases (HATs) have been well characterized both structurally and mechanistically, far less is known about non-histone acetyltransferase enzymes. The human ESCO1 and ESCO2 paralogs acetylate the cohesin complex subunit SMC3 to regulate the separation of sister chromatids during mitosis and meiosis. Missense mutations within the acetyltransferase domain of these proteins correlate with diseases, including endometrial cancers and Roberts syndrome. Despite their biological importance, the mechanisms underlying acetylation by the ESCO proteins are not understood. Here, we report the X-ray crystal structure of the highly conserved zinc finger-acetyltransferase moiety of ESCO1 with accompanying structure-based mutagenesis and biochemical characterization. We find that the ESCO1 acetyltransferase core is structurally homologous to the Gcn5 HAT, but contains unique additional features including a zinc finger and an ∼40-residue loop region that appear to play roles in protein stability and SMC3 substrate binding. We identify key residues that play roles in substrate binding and catalysis, and rationalize the functional consequences of disease-associated mutations. Together, these studies reveal the molecular basis for SMC3 acetylation by ESCO1 and have broader implications for understanding the structure/function of non-histone acetyltransferases.
Keywords: ESCO1; SMC3; X-ray crystallography; acetyltransferase; cell division; chromatin regulation; cohesin; crystal structure; enzyme structure; gene expression; posttranslational modification (PTM); protein acylation; protein structure; structure-function.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

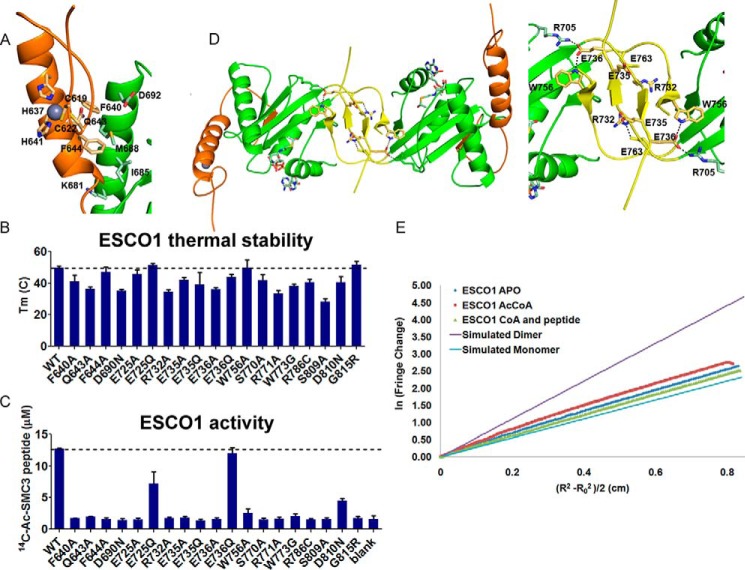


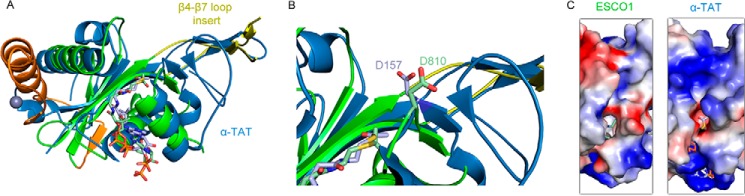
References
-
- Choudhary C., Kumar C., Gnad F., Nielsen M. L., Rehman M., Walther T. C., Olsen J. V., and Mann M. (2009) Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science 325, 834–840 - PubMed
-
- Yang X. J., and Seto E. (2007) HATs and HDACs: from structure, function and regulation to novel strategies for therapy and prevention. Oncogene 26, 5310–5318 - PubMed
-
- Tanner K. G., Langer M. R., and Denu J. M. (2000) Kinetic mechanism of human histone acetyltransferase P/CAF. Biochemistry 39, 11961–11969; Correction Biochemistry (2000) 39, 15652 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
