

Rapid and independent memory formation in the parietal cortex
- PMID: 27803331
- PMCID: PMC5135314
- DOI: 10.1073/pnas.1605719113
Rapid and independent memory formation in the parietal cortex
Abstract
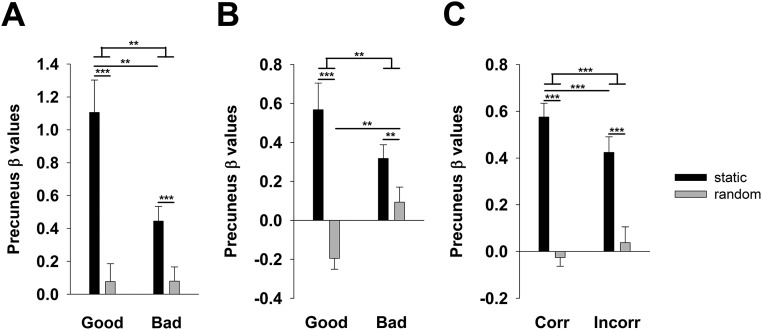
Previous evidence indicates that the brain stores memory in two complementary systems, allowing both rapid plasticity and stable representations at different sites. For memory to be established in a long-lasting neocortical store, many learning repetitions are considered necessary after initial encoding into hippocampal circuits. To elucidate the dynamics of hippocampal and neocortical contributions to the early phases of memory formation, we closely followed changes in human functional brain activity while volunteers navigated through two different, initially unknown virtual environments. In one condition, they were able to encode new information continuously about the spatial layout of the maze. In the control condition, no information could be learned because the layout changed constantly. Our results show that the posterior parietal cortex (PPC) encodes memories for spatial locations rapidly, beginning already with the first visit to a location and steadily increasing activity with each additional encounter. Hippocampal activity and connectivity between the PPC and hippocampus, on the other hand, are strongest during initial encoding, and both decline with additional encounters. Importantly, stronger PPC activity related to higher memory-based performance. Compared with the nonlearnable control condition, PPC activity in the learned environment remained elevated after a 24-h interval, indicating a stable change. Our findings reflect the rapid creation of a memory representation in the PPC, which belongs to a recently proposed parietal memory network. The emerging parietal representation is specific for individual episodes of experience, predicts behavior, and remains stable over offline periods, and must therefore hold a mnemonic function.
Keywords: long-term memory; memory systems consolidation; posterior parietal cortex; precuneus; virtual reality.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
-
- Frankland PW, Bontempi B. The organization of recent and remote memories. Nat Rev Neurosci. 2005;6(2):119–130. - PubMed
-
- McClelland JL, McNaughton BL, O’Reilly RC. Why there are complementary learning systems in the hippocampus and neocortex: Insights from the successes and failures of connectionist models of learning and memory. Psychol Rev. 1995;102(3):419–457. - PubMed
-
- Squire LR, Zola-Morgan S. The medial temporal lobe memory system. Science. 1991;253(5026):1380–1386. - PubMed
-
- Marr D. A theory for cerebral neocortex. Proc R Soc Lond B Biol Sci. 1970;176(1043):161–234. - PubMed
-
- Maviel T, Durkin TP, Menzaghi F, Bontempi B. Sites of neocortical reorganization critical for remote spatial memory. Science. 2004;305(5680):96–99. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
