

Virus-Infected Human Mast Cells Enhance Natural Killer Cell Functions
- PMID: 27806369
- PMCID: PMC6738812
- DOI: 10.1159/000450576
Virus-Infected Human Mast Cells Enhance Natural Killer Cell Functions
Abstract
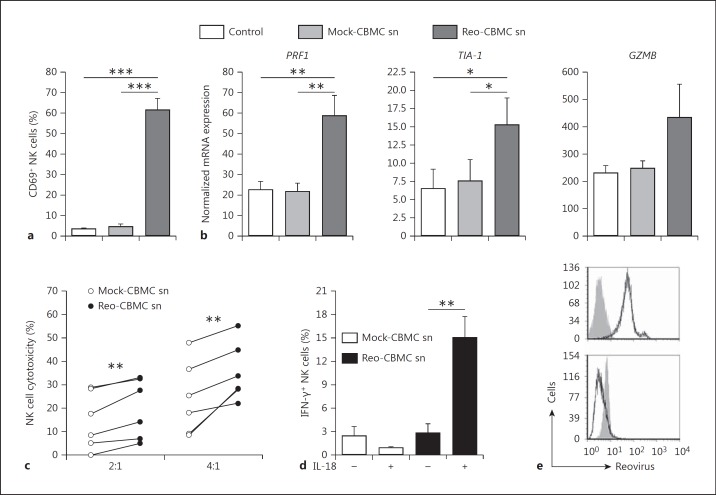
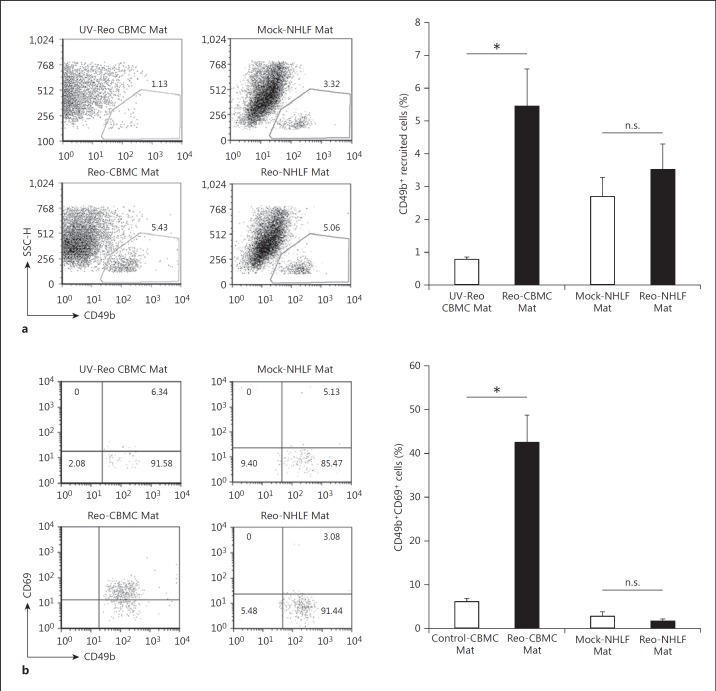
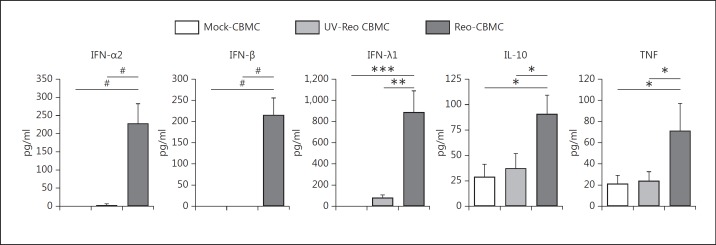
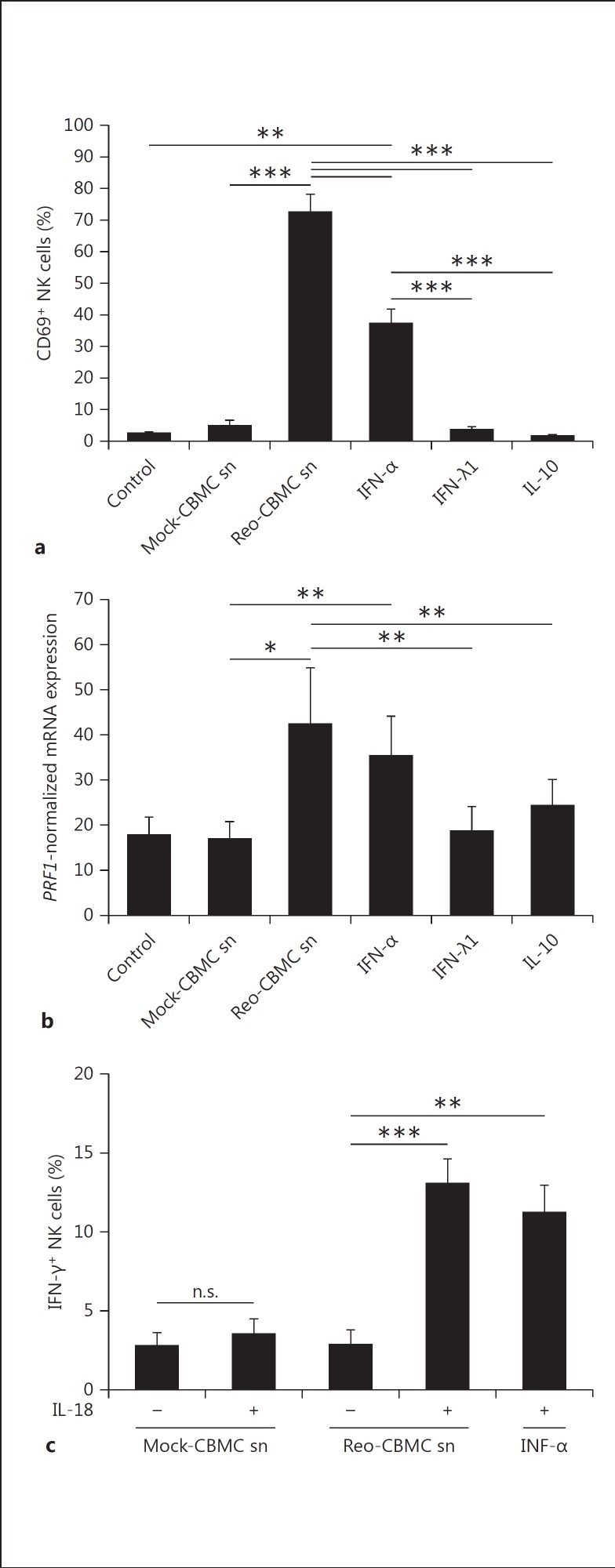
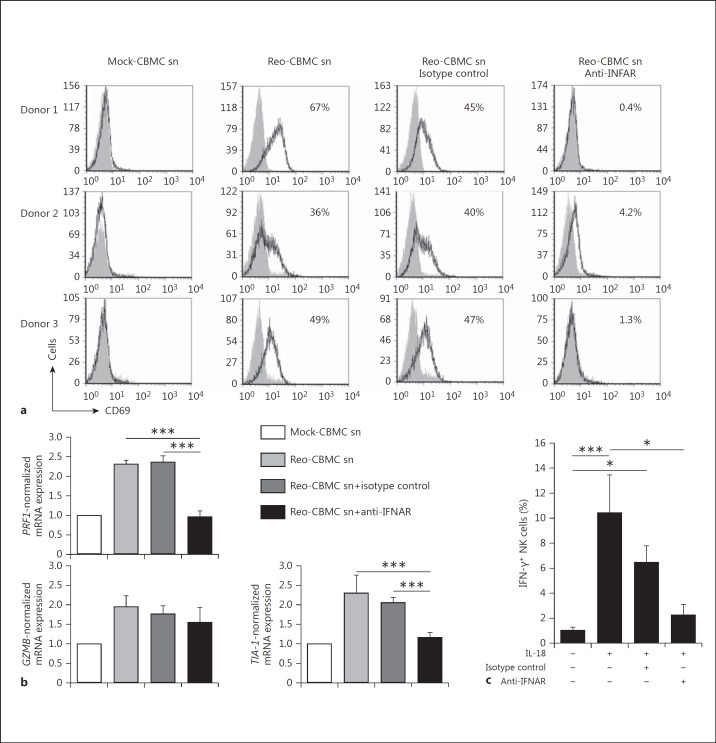
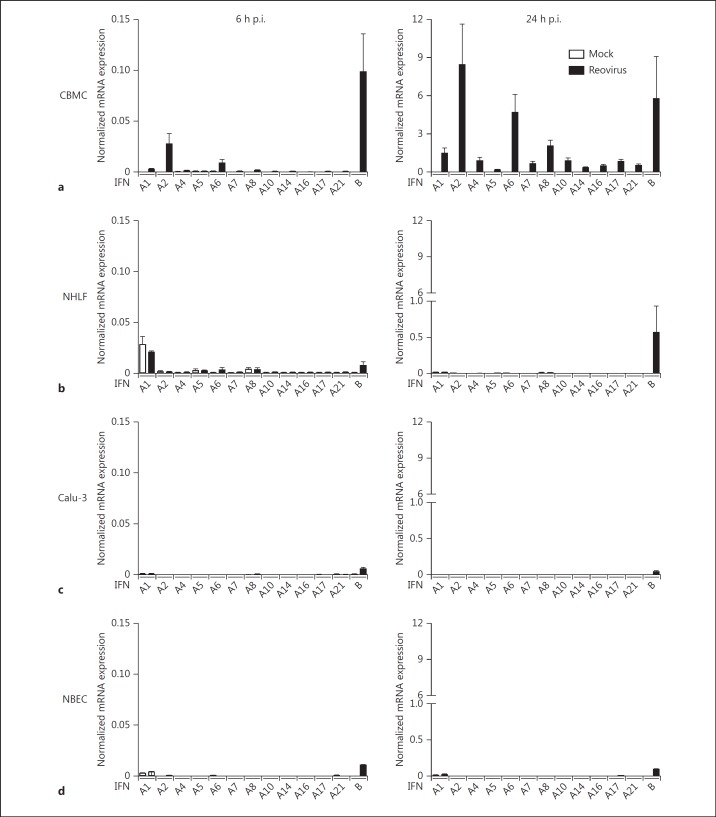
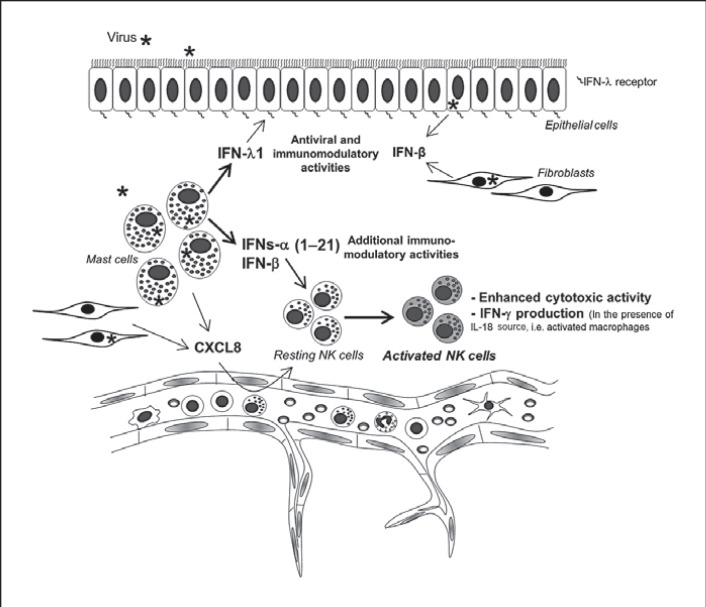
Mucosal surfaces are protected from infection by both structural and sentinel cells, such as mast cells. The mast cell's role in antiviral responses is poorly understood; however, they selectively recruit natural killer (NK) cells following infection. Here, the ability of virus-infected mast cells to enhance NK cell functions was examined. Cord blood-derived human mast cells infected with reovirus (Reo-CBMC) and subsequent mast cell products were used for the stimulation of human NK cells. NK cells upregulated the CD69 molecule and cytotoxicity-related genes, and demonstrated increased cytotoxic activity in response to Reo-CBMC soluble products. NK cell interferon (IFN)-γ production was also promoted in the presence of interleukin (IL)-18. In vivo, SCID mice injected with Reo-CBMC in a subcutaneous Matrigel model, could recruit and activate murine NK cells, a property not shared by normal human fibroblasts. Soluble products of Reo-CBMC included IL-10, TNF, type I and type III IFNs. Blockade of the type I IFN receptor abrogated NK cell activation. Furthermore, reovirus-infected mast cells expressed multiple IFN-α subtypes not observed in reovirus-infected fibroblasts or epithelial cells. Our data define an important mast cell IFN response, not shared by structural cells, and a subsequent novel mast cell-NK cell immune axis in human antiviral host defense.
© 2016 S. Karger AG, Basel.
Figures
Similar articles
-
Mast Cells and Natural Killer Cells-A Potentially Critical Interaction.Viruses. 2019 Jun 4;11(6):514. doi: 10.3390/v11060514. Viruses. 2019. PMID: 31167464 Free PMC article. Review.
-
Human mast cell activation with virus-associated stimuli leads to the selective chemotaxis of natural killer cells by a CXCL8-dependent mechanism.Blood. 2008 Jun 15;111(12):5467-76. doi: 10.1182/blood-2007-10-118547. Epub 2008 Apr 18. Blood. 2008. PMID: 18424663
-
Virus stimulation of human mast cells results in the recruitment of CD56⁺ T cells by a mechanism dependent on CCR5 ligands.FASEB J. 2012 Mar;26(3):1280-9. doi: 10.1096/fj.11-188979. Epub 2011 Nov 28. FASEB J. 2012. PMID: 22125314
-
Reciprocal human dendritic cell-natural killer cell interactions induce antitumor activity following tumor cell infection by oncolytic reovirus.J Immunol. 2009 Oct 1;183(7):4312-21. doi: 10.4049/jimmunol.0901074. Epub 2009 Sep 4. J Immunol. 2009. PMID: 19734207
-
Interferon--a major regulator of natural killer cell-mediated cytotoxicity.J Interferon Res. 1993 Aug;13(4):247-57. doi: 10.1089/jir.1993.13.247. J Interferon Res. 1993. PMID: 7693829 Review.
Cited by
-
Mast Cell Activation Triggered by Retrovirus Promotes Acute Viral Infection.Front Microbiol. 2022 Feb 7;13:798660. doi: 10.3389/fmicb.2022.798660. eCollection 2022. Front Microbiol. 2022. PMID: 35197951 Free PMC article.
-
Mast Cells and Natural Killer Cells-A Potentially Critical Interaction.Viruses. 2019 Jun 4;11(6):514. doi: 10.3390/v11060514. Viruses. 2019. PMID: 31167464 Free PMC article. Review.
-
Protective and pathogenic roles for mast cells during viral infections.Curr Opin Immunol. 2020 Oct;66:74-81. doi: 10.1016/j.coi.2020.05.003. Epub 2020 Jun 18. Curr Opin Immunol. 2020. PMID: 32563779 Free PMC article. Review.
-
Steering Mast Cells or Their Mediators as a Prospective Novel Therapeutic Approach for the Treatment of Hematological Malignancies.Front Oncol. 2021 Sep 24;11:731323. doi: 10.3389/fonc.2021.731323. eCollection 2021. Front Oncol. 2021. PMID: 34631562 Free PMC article. Review.
-
Crosstalk of Mast Cells and Natural Killer Cells with Neurons in Chemotherapy-Induced Peripheral Neuropathy.Int J Mol Sci. 2023 Aug 8;24(16):12543. doi: 10.3390/ijms241612543. Int J Mol Sci. 2023. PMID: 37628724 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
