

Hair follicles' transit-amplifying cells govern concurrent dermal adipocyte production through Sonic Hedgehog
- PMID: 27807033
- PMCID: PMC5110998
- DOI: 10.1101/gad.285429.116
Hair follicles' transit-amplifying cells govern concurrent dermal adipocyte production through Sonic Hedgehog
Abstract
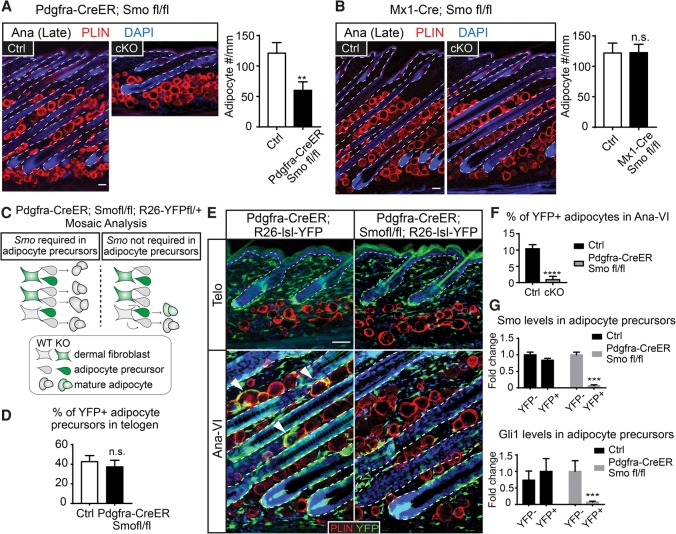
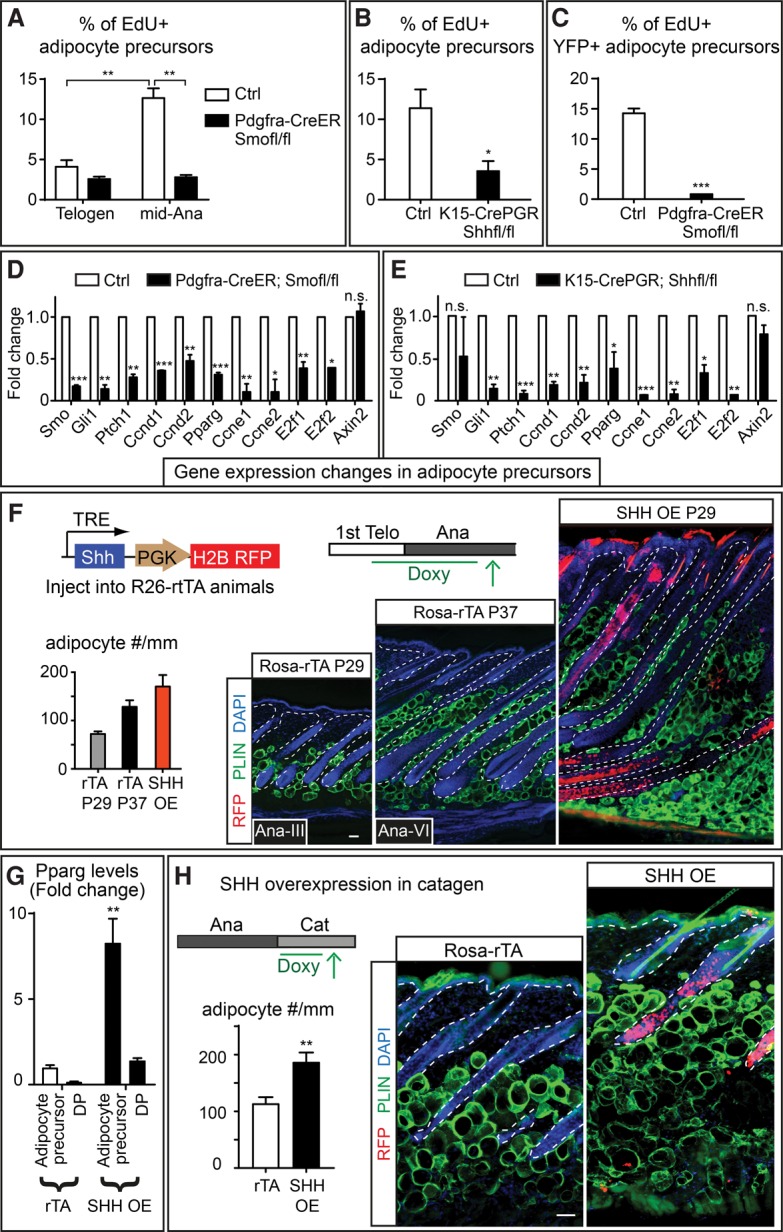
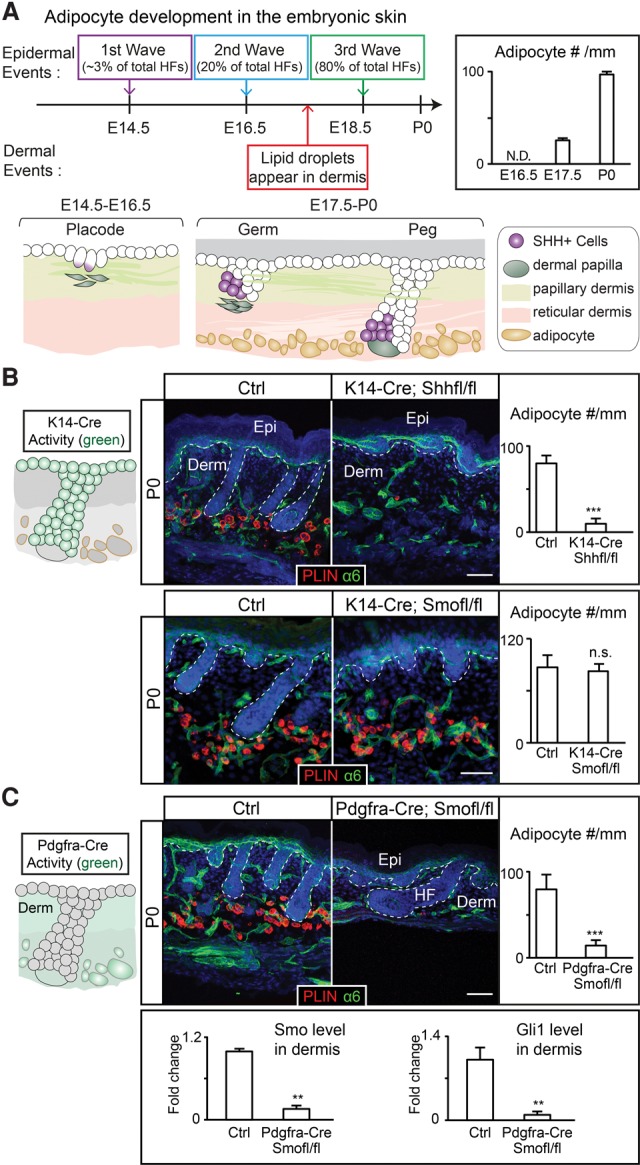
Growth and regeneration of one tissue within an organ compels accommodative changes in the surrounding tissues. However, the molecular nature and operating logic governing these concurrent changes remain poorly defined. The dermal adipose layer expands concomitantly with hair follicle downgrowth, providing a paradigm for studying coordinated changes of surrounding lineages with a regenerating tissue. Here, we discover that hair follicle transit-amplifying cells (HF-TACs) play an essential role in orchestrating dermal adipogenesis through secreting Sonic Hedgehog (SHH). Depletion of Shh from HF-TACs abrogates both dermal adipogenesis and hair follicle growth. Using cell type-specific deletion of Smo, a gene required in SHH-receiving cells, we found that SHH does not act on hair follicles, adipocytes, endothelial cells, and hematopoietic cells for adipogenesis. Instead, SHH acts directly on adipocyte precursors, promoting their proliferation and their expression of a key adipogenic gene, peroxisome proliferator-activated receptor γ (Pparg), to induce dermal adipogenesis. Our study therefore uncovers a critical role for TACs in orchestrating the generation of both their own progeny and a neighboring lineage to achieve concomitant tissue production across lineages.
Keywords: adipocyte precursors; adipogenesis; hair follicle regeneration; interlineage communications; stroma; transit-amplifying cells.
© 2016 Zhang et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
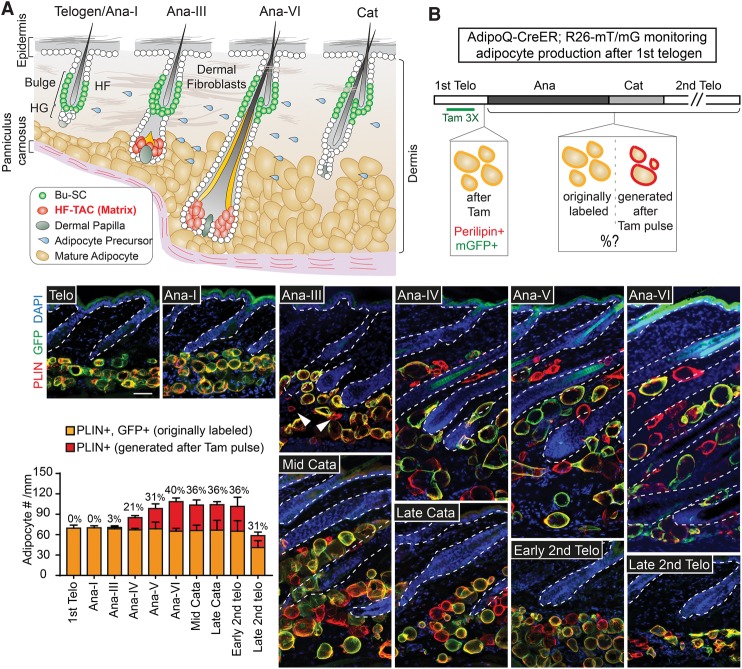
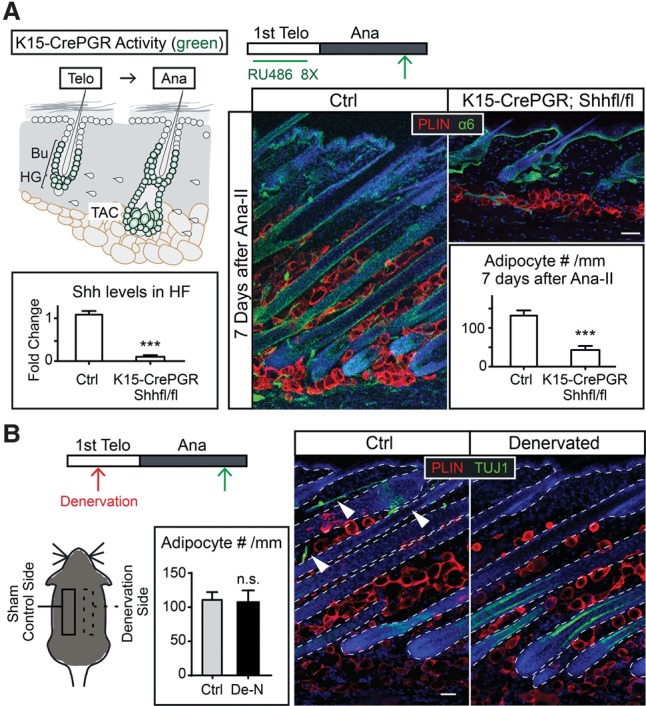
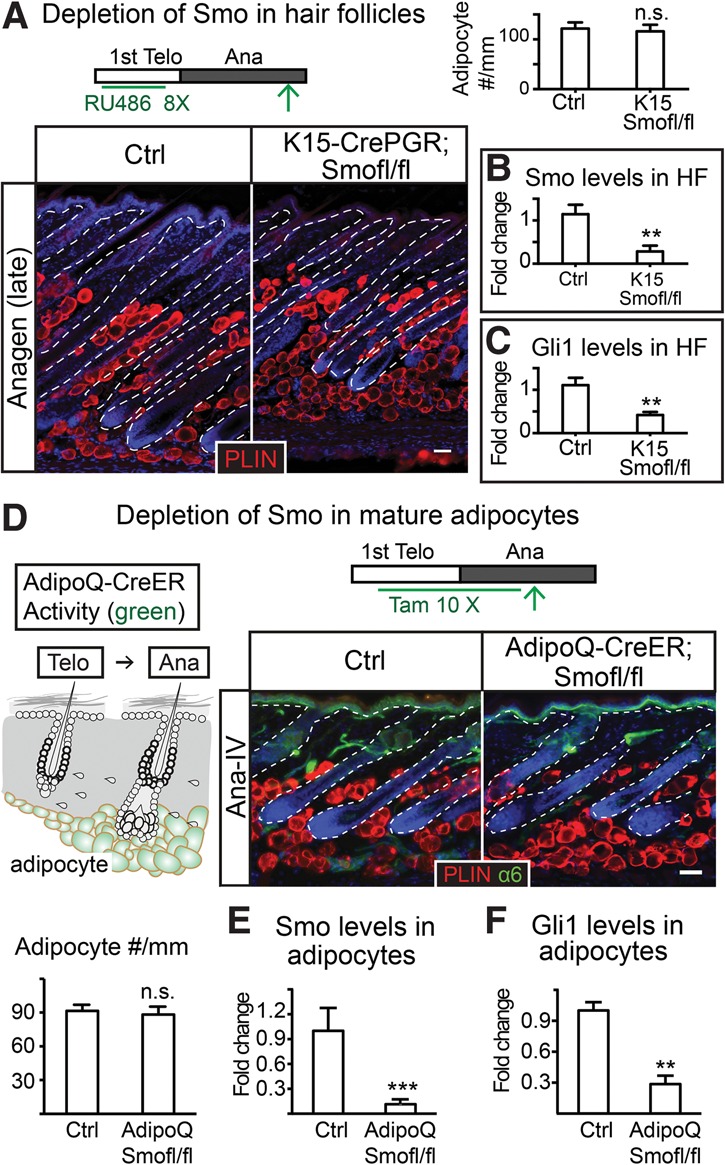
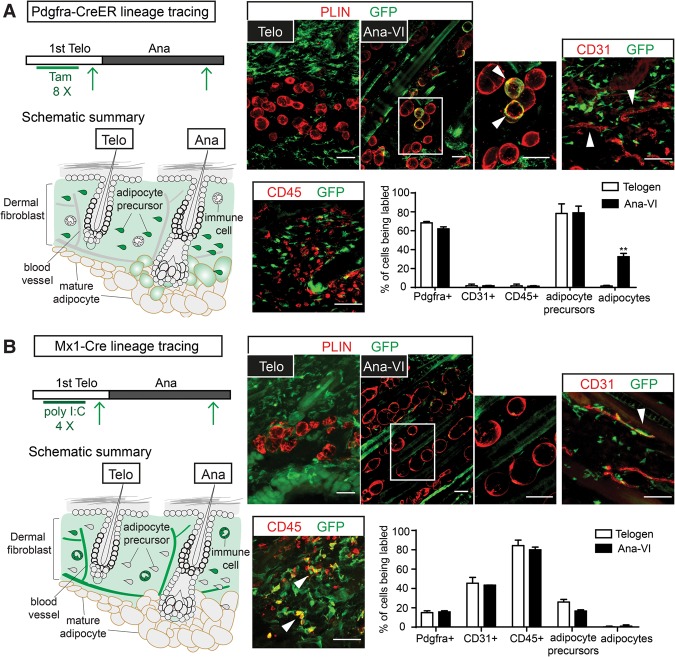
Comment in
-
Adipose Tissue: SHH and dermal adipogenesis.Nat Rev Endocrinol. 2017 Jan;13(1):4. doi: 10.1038/nrendo.2016.192. Epub 2016 Nov 18. Nat Rev Endocrinol. 2017. PMID: 27857128 No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous