

Emergence of the Noncoding Cancer Genome: A Target of Genetic and Epigenetic Alterations
- PMID: 27807102
- PMCID: PMC5117653
- DOI: 10.1158/2159-8290.CD-16-0745
Emergence of the Noncoding Cancer Genome: A Target of Genetic and Epigenetic Alterations
Abstract
The emergence of whole-genome annotation approaches is paving the way for the comprehensive annotation of the human genome across diverse cell and tissue types exposed to various environmental conditions. This has already unmasked the positions of thousands of functional cis-regulatory elements integral to transcriptional regulation, such as enhancers, promoters, and anchors of chromatin interactions that populate the noncoding genome. Recent studies have shown that cis-regulatory elements are commonly the targets of genetic and epigenetic alterations associated with aberrant gene expression in cancer. Here, we review these findings to showcase the contribution of the noncoding genome and its alteration in the development and progression of cancer. We also highlight the opportunities to translate the biological characterization of genetic and epigenetic alterations in the noncoding cancer genome into novel approaches to treat or monitor disease.
Significance: The majority of genetic and epigenetic alterations accumulate in the noncoding genome throughout oncogenesis. Discriminating driver from passenger events is a challenge that holds great promise to improve our understanding of the etiology of different cancer types. Advancing our understanding of the noncoding cancer genome may thus identify new therapeutic opportunities and accelerate our capacity to find improved biomarkers to monitor various stages of cancer development. Cancer Discov; 6(11); 1215-29. ©2016 AACR.
©2016 American Association for Cancer Research.
Conflict of interest statement
of Potential Conflicts of Interest No potential conflicts of interest to disclose.
Figures
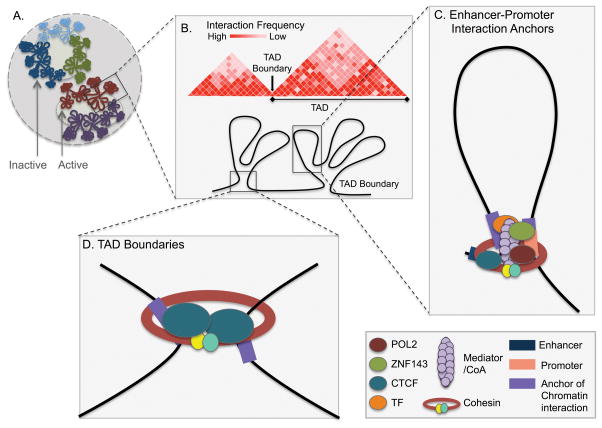
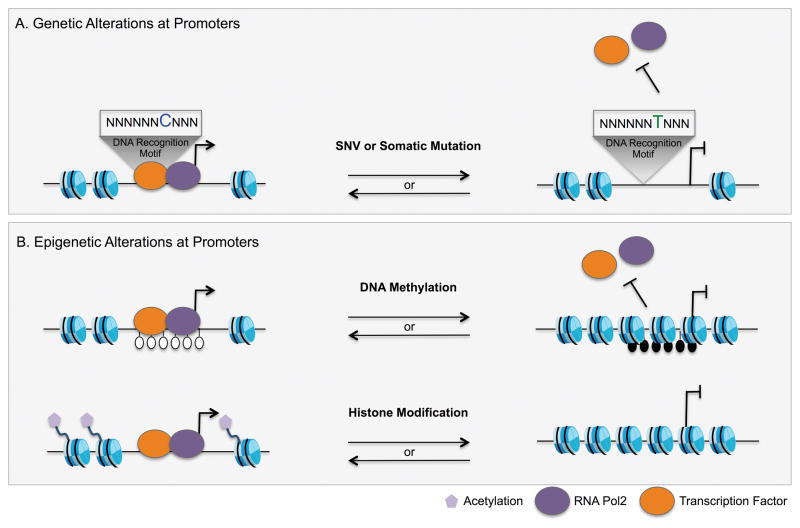
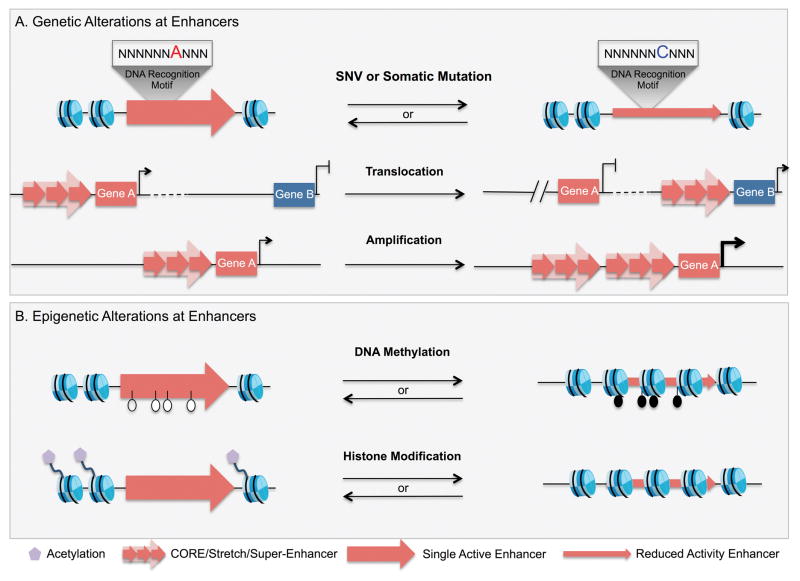
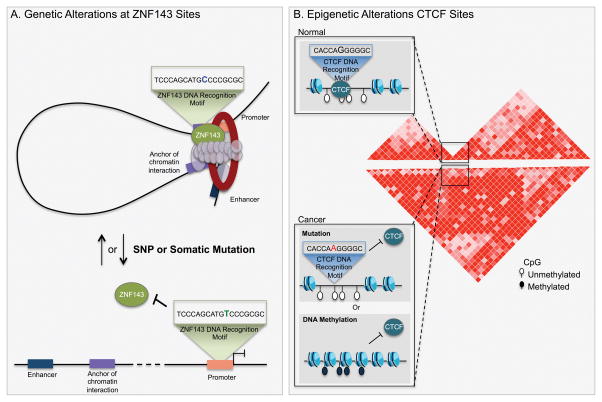
References
-
- Wittkopp PJ, Kalay G. Cis-regulatory elements: molecular mechanisms and evolutionary processes underlying divergence. Nat Rev Genet. 2012;13:59–69. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
