

The correlations and sequence of plant stomatal, hydraulic, and wilting responses to drought
- PMID: 27807136
- PMCID: PMC5135344
- DOI: 10.1073/pnas.1604088113
The correlations and sequence of plant stomatal, hydraulic, and wilting responses to drought
Abstract
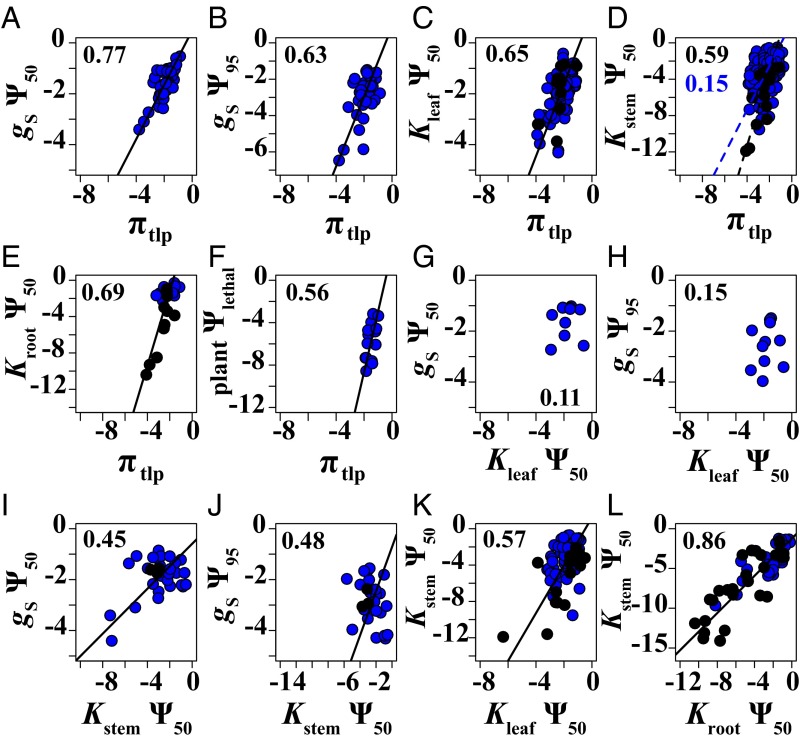
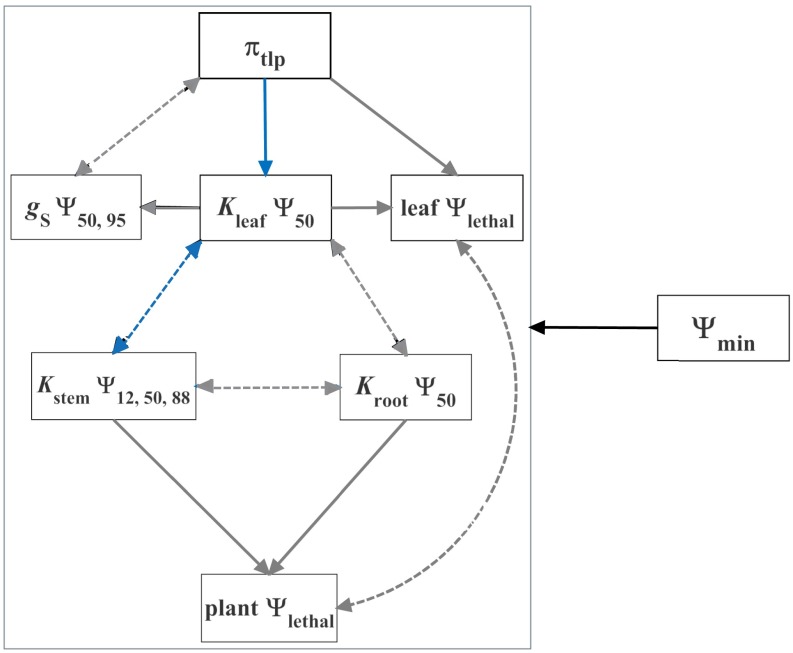
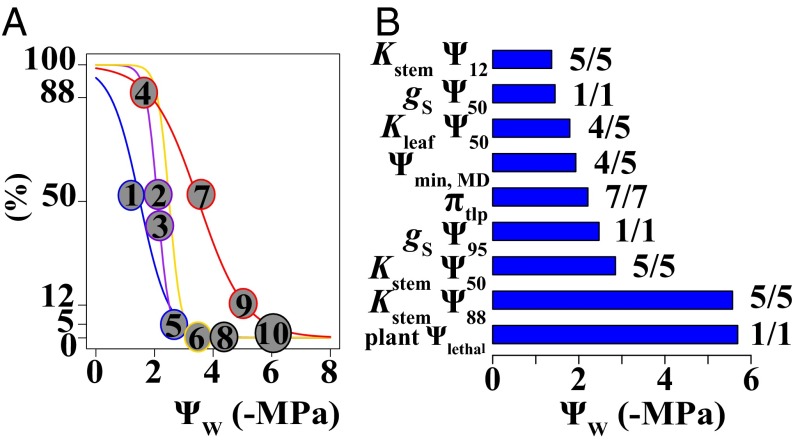
Climate change is expected to exacerbate drought for many plants, making drought tolerance a key driver of species and ecosystem responses. Plant drought tolerance is determined by multiple traits, but the relationships among traits, either within individual plants or across species, have not been evaluated for general patterns across plant diversity. We synthesized the published data for stomatal closure, wilting, declines in hydraulic conductivity in the leaves, stems, and roots, and plant mortality for 262 woody angiosperm and 48 gymnosperm species. We evaluated the correlations among the drought tolerance traits across species, and the general sequence of water potential thresholds for these traits within individual plants. The trait correlations across species provide a framework for predicting plant responses to a wide range of water stress from one or two sampled traits, increasing the ability to rapidly characterize drought tolerance across diverse species. Analyzing these correlations also identified correlations among the leaf and stem hydraulic traits and the wilting point, or turgor loss point, beyond those expected from shared ancestry or independent associations with water stress alone. Further, on average, the angiosperm species generally exhibited a sequence of drought tolerance traits that is expected to limit severe tissue damage during drought, such as wilting and substantial stem embolism. This synthesis of the relationships among the drought tolerance traits provides crucial, empirically supported insight into representing variation in multiple traits in models of plant and ecosystem responses to drought.
Keywords: drought tolerance; leaf hydraulics; stem hydraulics; stomatal closure; turgor loss point.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Sheffield J, Wood EF. Characteristics of global and regional drought, 1950–2000: Analysis of soil moisture data from off-line simulation of the terrestrial hydrologic cycle. J Geophys Res. 2007;112(D17):115.
-
- Maherali H, Pockman WT, Jackson RB. Adaptive variation in the vulnerability of woody plants to xylem cavitation. Ecology. 2004;85(8):2184–2199.
-
- Choat B, et al. Global convergence in the vulnerability of forests to drought. Nature. 2012;491(7426):752–755. - PubMed
-
- Blackman CJ, Brodribb TJ, Jordan GJ. Leaf hydraulic vulnerability influences species’ bioclimatic limits in a diverse group of woody angiosperms. Oecologia. 2012;168(1):1–10. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
