

Estrogens and Androgens in Skeletal Physiology and Pathophysiology
- PMID: 27807202
- PMCID: PMC5539371
- DOI: 10.1152/physrev.00033.2015
Estrogens and Androgens in Skeletal Physiology and Pathophysiology
Abstract
Estrogens and androgens influence the growth and maintenance of the mammalian skeleton and are responsible for its sexual dimorphism. Estrogen deficiency at menopause or loss of both estrogens and androgens in elderly men contribute to the development of osteoporosis, one of the most common and impactful metabolic diseases of old age. In the last 20 years, basic and clinical research advances, genetic insights from humans and rodents, and newer imaging technologies have changed considerably the landscape of our understanding of bone biology as well as the relationship between sex steroids and the physiology and pathophysiology of bone metabolism. Together with the appreciation of the side effects of estrogen-related therapies on breast cancer and cardiovascular diseases, these advances have also drastically altered the treatment of osteoporosis. In this article, we provide a comprehensive review of the molecular and cellular mechanisms of action of estrogens and androgens on bone, their influences on skeletal homeostasis during growth and adulthood, the pathogenetic mechanisms of the adverse effects of their deficiency on the female and male skeleton, as well as the role of natural and synthetic estrogenic or androgenic compounds in the pharmacotherapy of osteoporosis. We highlight latest advances on the crosstalk between hormonal and mechanical signals, the relevance of the antioxidant properties of estrogens and androgens, the difference of their cellular targets in different bone envelopes, the role of estrogen deficiency in male osteoporosis, and the contribution of estrogen or androgen deficiency to the monomorphic effects of aging on skeletal involution.
Copyright © 2017 the American Physiological Society.
Figures
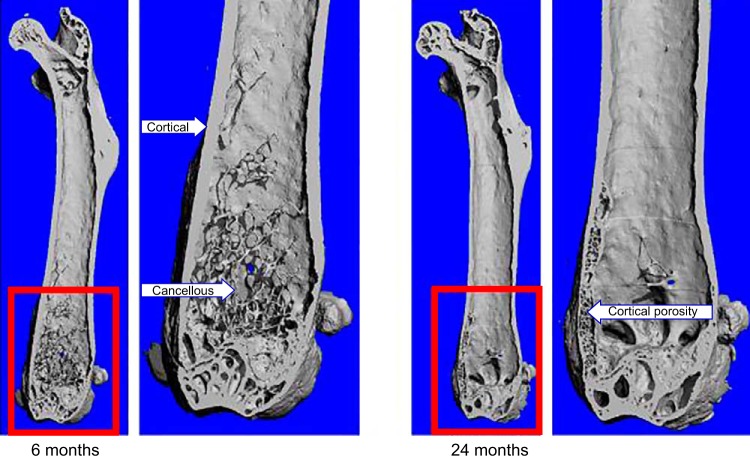
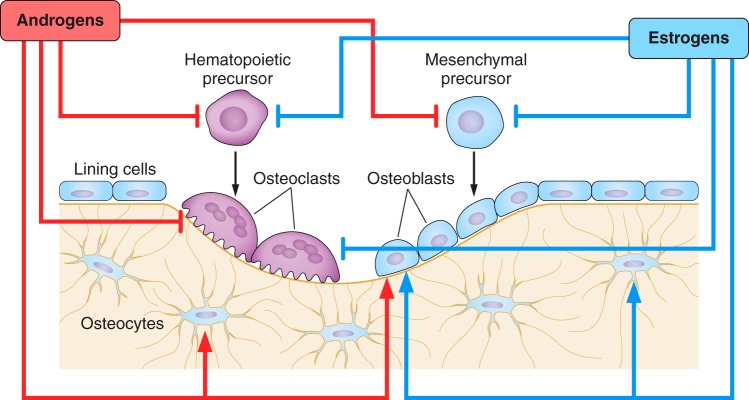
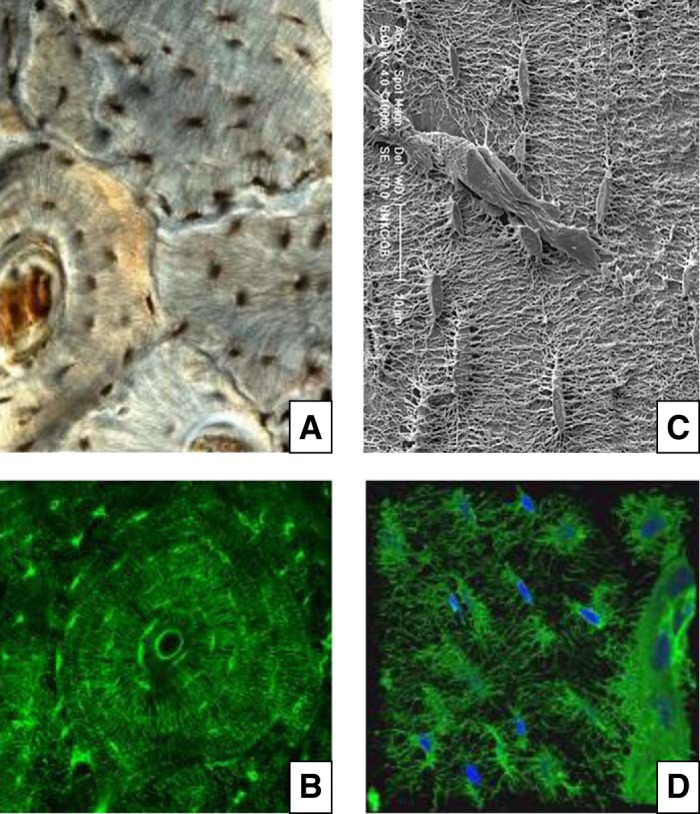
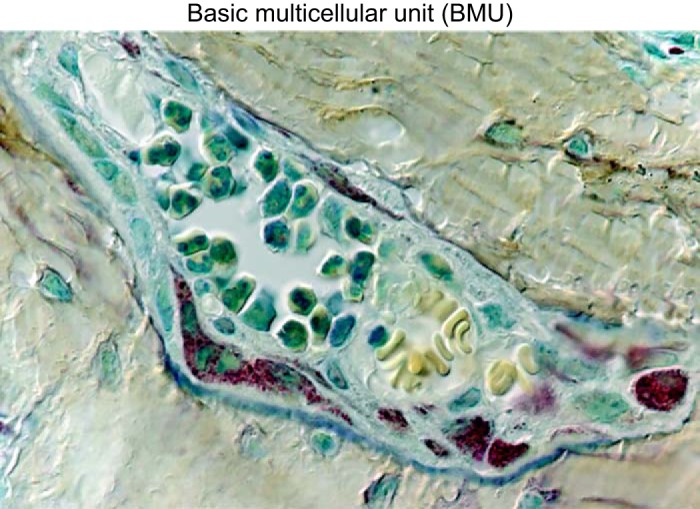
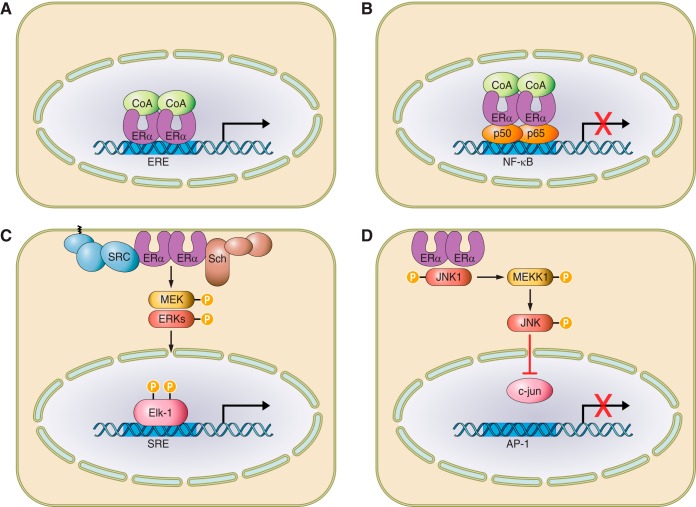
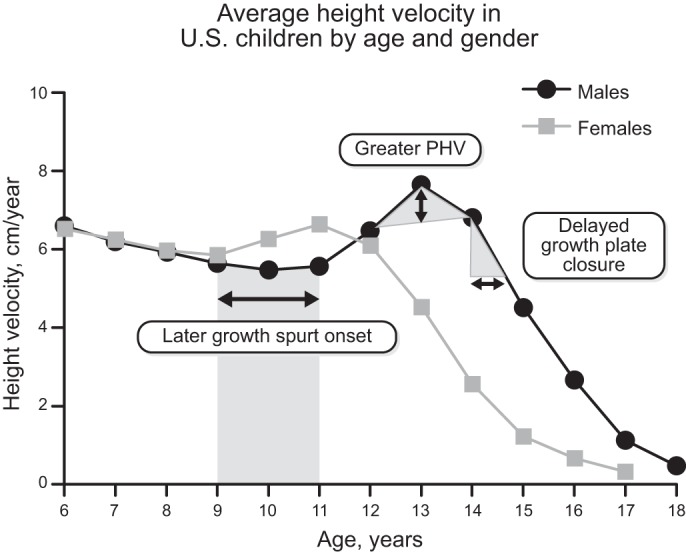
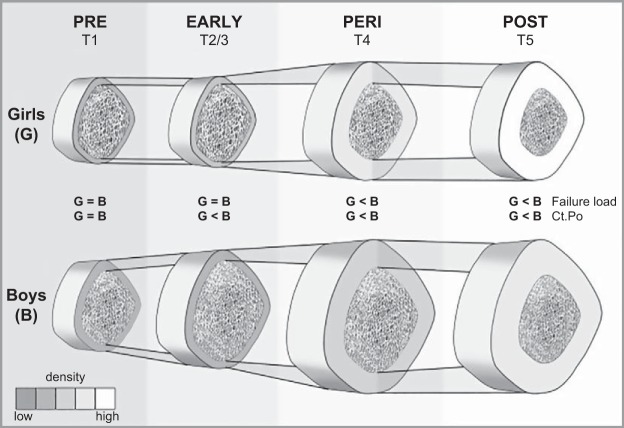
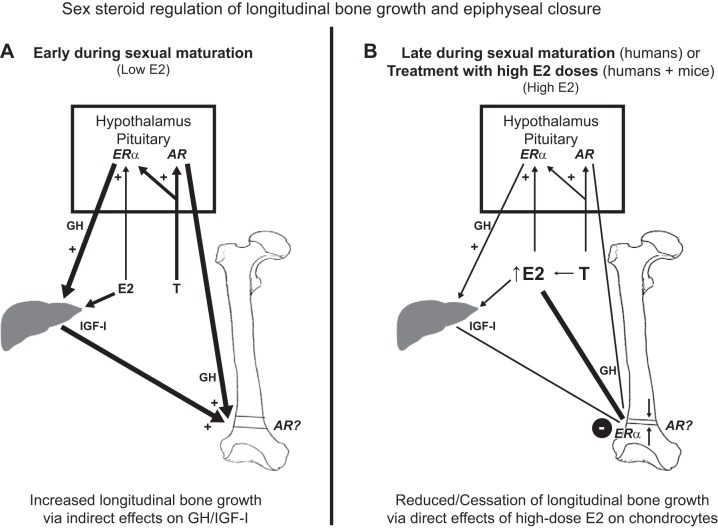
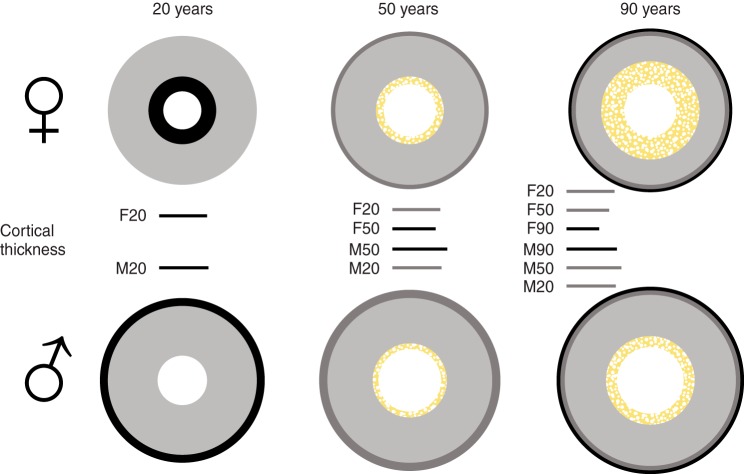
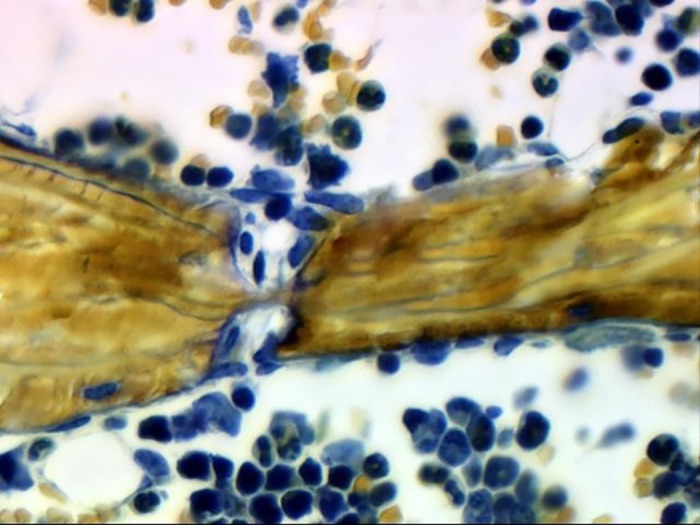
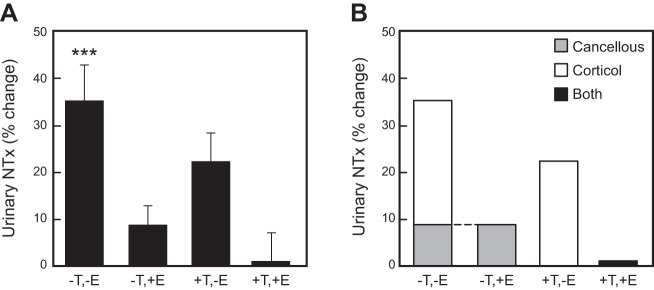
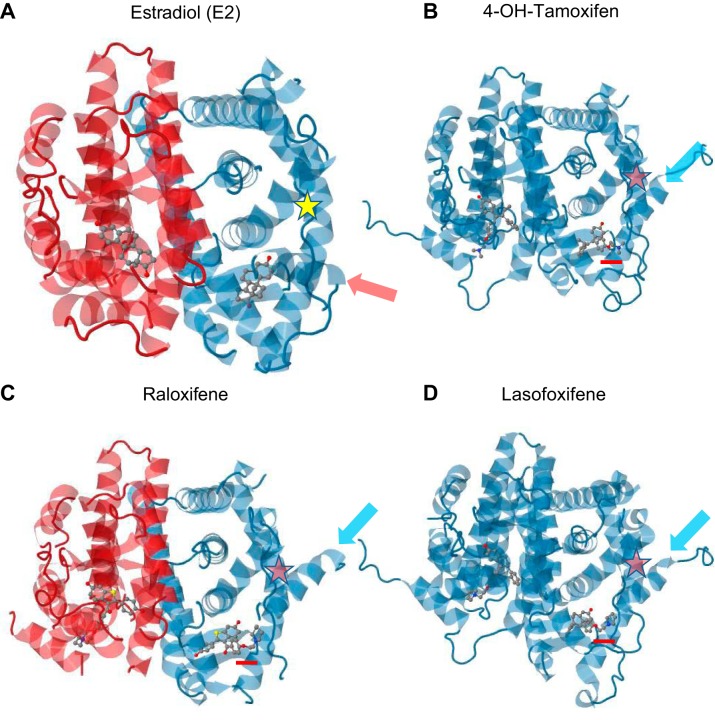
References
-
- National Institute of Health Consensus Development Panel on Osteoporosis Prevention, Diagnosis, and Therapy. JAMA 285: 785–795, 2001. - PubMed
-
- Osteoporosis in Men: The Effects of Gender on Skeletal Health. London: Elsevier, 2010.
-
- Aarden EM, Burger EH, Nijweide PJ. Function of osteocytes in bone. J Cell Biochem 55: 287–299, 1994. - PubMed
-
- Adams GB, Scadden DT. The hematopoietic stem cell in its place. Nat Immunol 7: 333–337, 2006. - PubMed
-
- Aguirre JI, Plotkin LI, Gortazar AR, Millan MM, O'Brien CA, Manolagas SC, Bellido T. A novel ligand-independent function of the estrogen receptor is essential for osteocyte and osteoblast mechanotransduction. J Biol Chem 282: 25501–25508, 2007. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical