

α-Synuclein enhances histone H3 lysine-9 dimethylation and H3K9me2-dependent transcriptional responses
- PMID: 27808254
- PMCID: PMC5093762
- DOI: 10.1038/srep36328
α-Synuclein enhances histone H3 lysine-9 dimethylation and H3K9me2-dependent transcriptional responses
Abstract
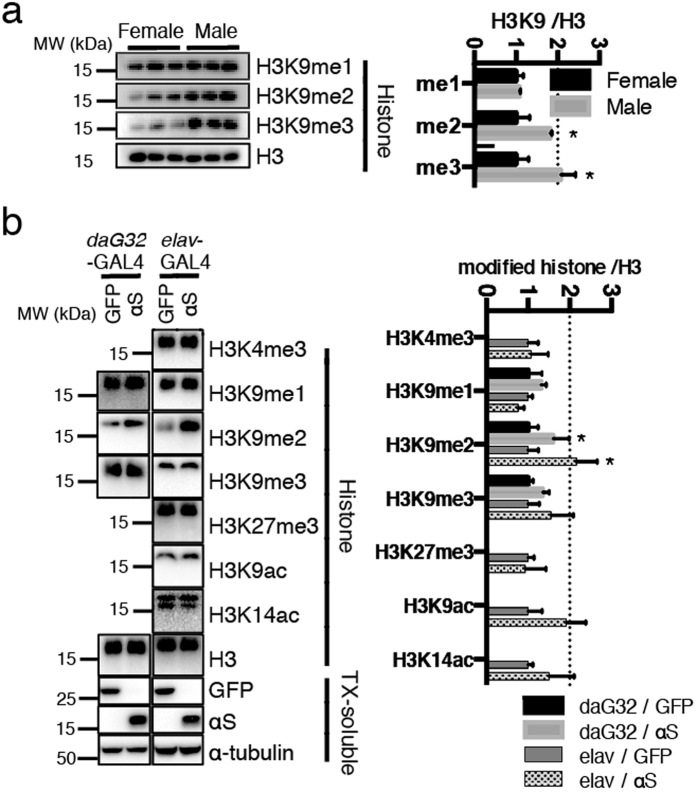
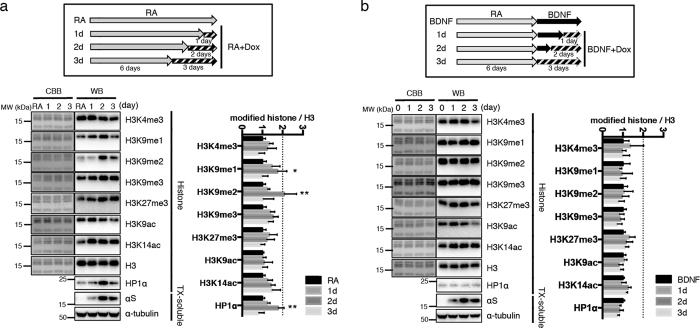
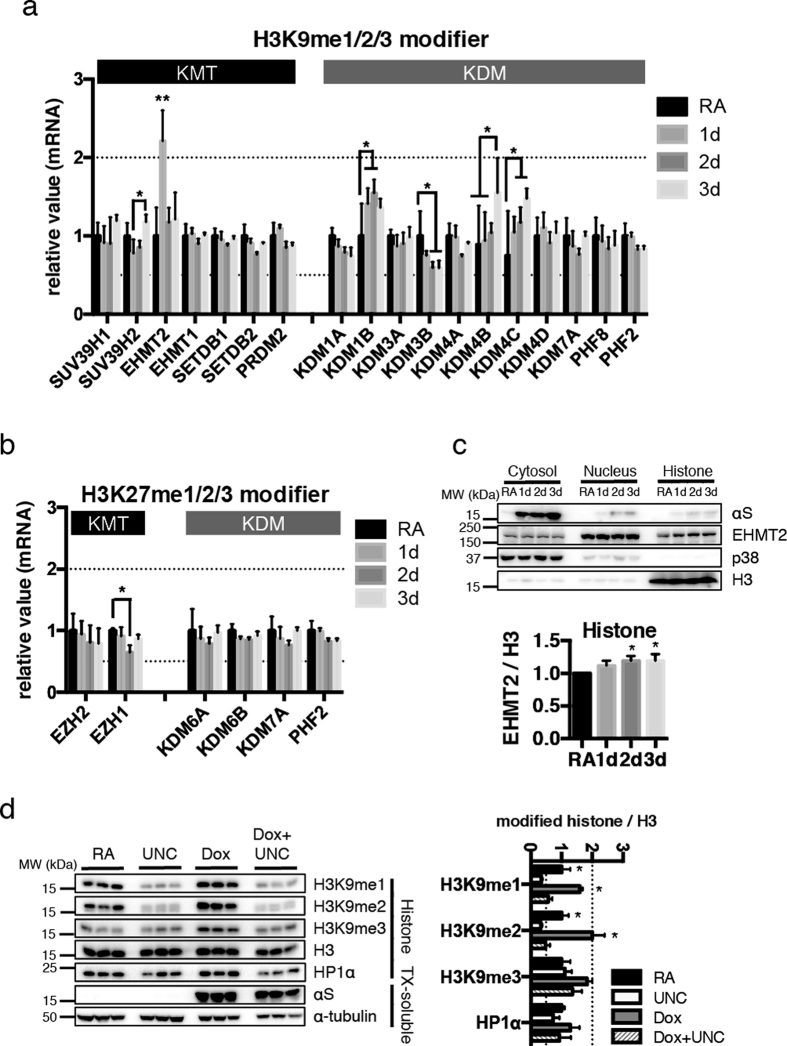
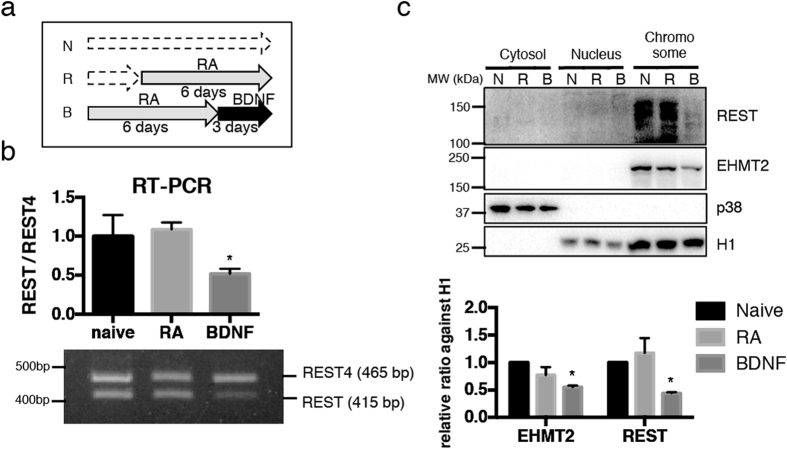
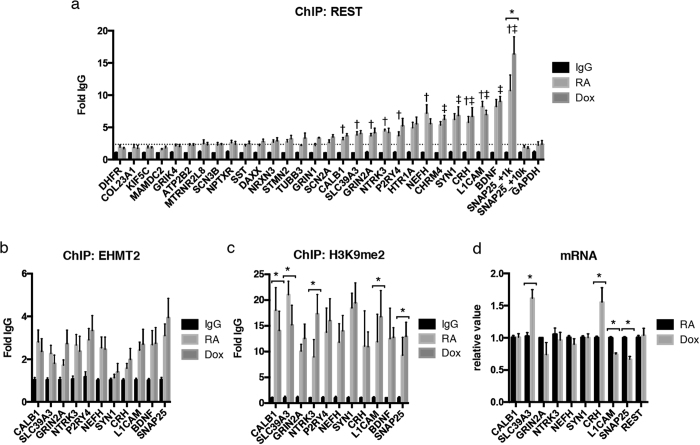
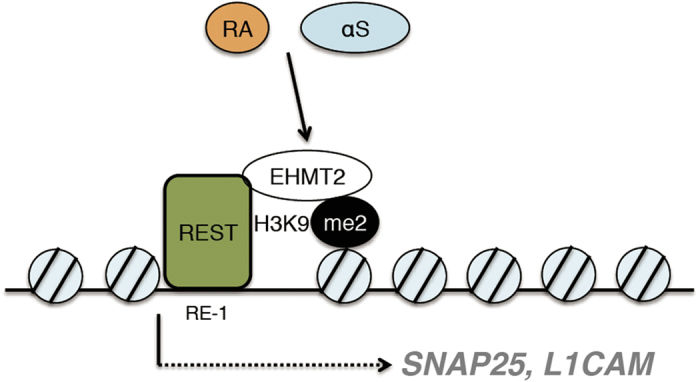
α-Synuclein (αS) is a protein linked to Parkinson's disease (PD) and related neurodegenerative disorders. It is mostly localized within synapses, but αS has also been suggested to play a role in the nucleus. We used transgenic Drosophila and inducible SH-SY5Y neuroblastoma cells to investigate the effects of αS on chromatin with a particular focus on histone modifications. Overexpression of αS in male flies as well as in retinoic acid pre-treated neuroblastoma cells led to an elevation of histone H3K9 methylations, mostly mono- (H3K9me1) and di- (H3K9me2). The transient increase of H3K9 methylation in αS-induced SH-SY5Y cells was preceded by mRNA induction of the euchromatic histone lysine N-methyltransferase 2 (EHMT2). EHMT2 and H3K9me2 can function within the REST complex. Chromatin immunoprecipitation (ChIP) analyses of selected candidate, REST regulated genes showed significantly increased H3K9me2 promoter occupancy of genes encoding the L1CAM cell adhesion molecule and the synaptosomal-associated protein SNAP25, whose reduced expression levels were confirmed by RT-qPCR in αS induced cells. Treatment with EHMT inhibitor UNC0638 restored the mRNA levels of L1CAM and SNAP25. Thus, αS overexpression enhances H3K9 methylations via ΕΗΜΤ2 resulting in elevated H3K9me2 at the SNAP25 promoter, possibly affecting SNARE complex assembly and hence synaptic vesicle fusion events regulated by αS.
Figures

References
-
- Polymeropoulos M. H. et al.. Mutation in the alpha-synuclein gene identified in families with Parkinson’s disease. Science 276, 2045–2047 (1997). - PubMed
-
- Eriksen J. L., Dawson T. M., Dickson D. W. & Petrucelli L. Caught in the act: alpha-synuclein is the culprit in Parkinson’s disease. Neuron 40, 453–456 (2003). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
