

Cholinergic modulation of the parafacial respiratory group
- PMID: 27808424
- PMCID: PMC5309363
- DOI: 10.1113/JP273012
Cholinergic modulation of the parafacial respiratory group
Abstract
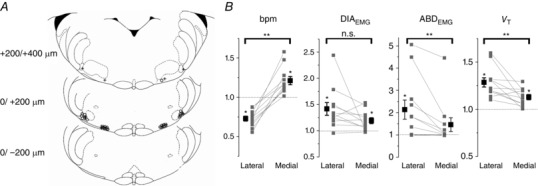
Key points: This study investigates the effects of cholinergic transmission on the expiratory oscillator, the parafacial respiratory group (pFRG) in urethane anaesthetized adult rats. Local inhibition of the acetyl cholinesterase enzyme induced activation of expiratory abdominal muscles and active expiration. Local application of the cholinomimetic carbachol elicited recruitment of late expiratory neurons, expiratory abdominal muscle activity and active expiration. This effect was antagonized by local application of the muscarinic antagonists scopolamine, J104129 and 4DAMP. We observed distinct physiological responses between the more medial chemosensitive region of the retrotrapezoid nucleus and the more lateral region of pFRG. These results support the hypothesis that pFRG is under cholinergic neuromodulation and the region surrounding the facial nucleus contains a group of neurons with distinct physiological roles.
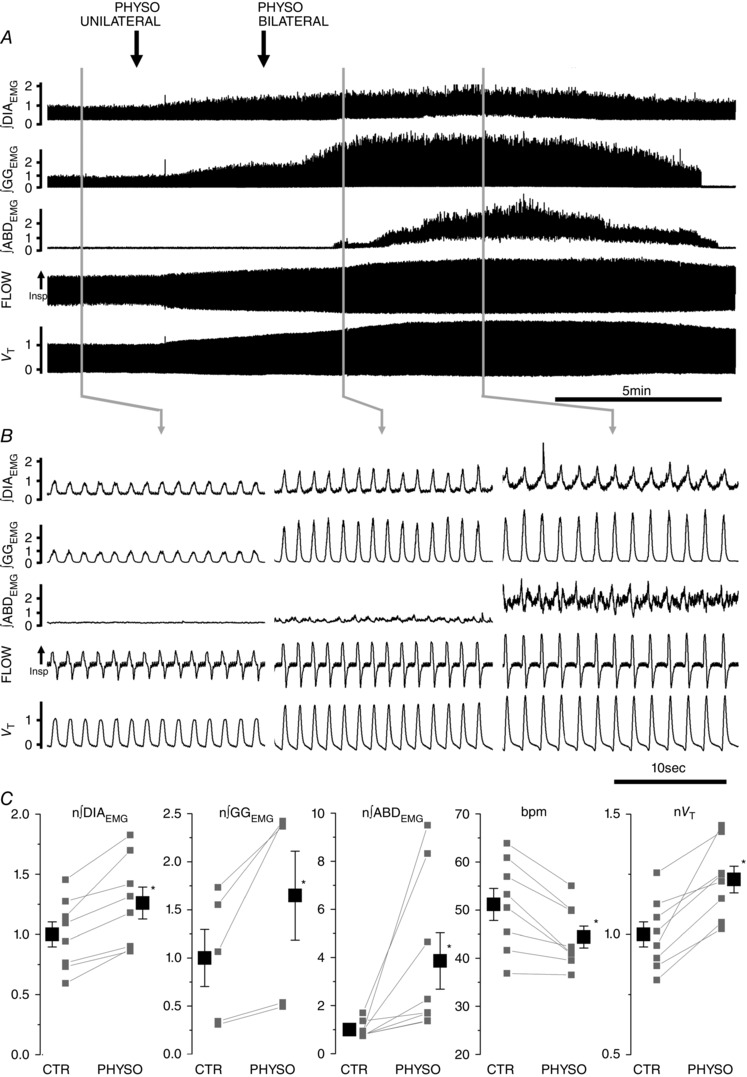
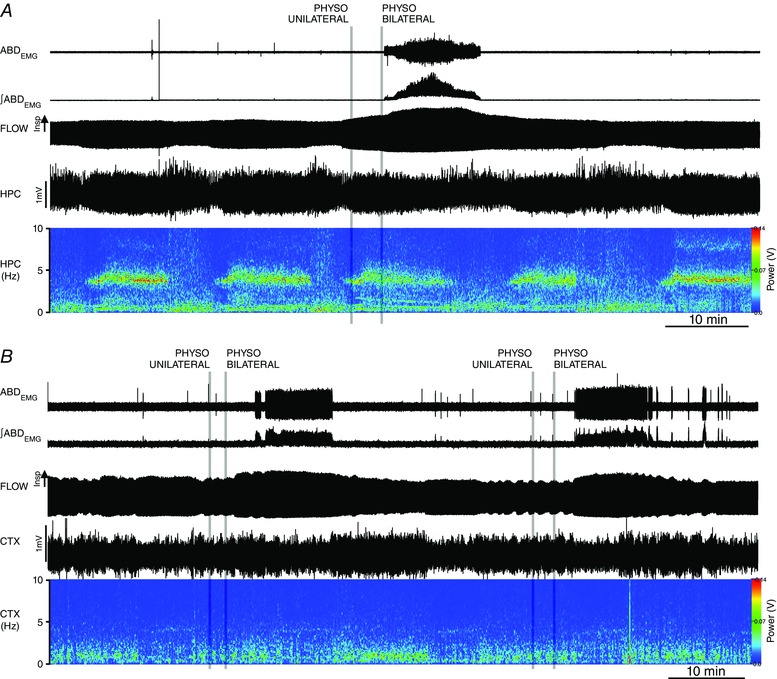
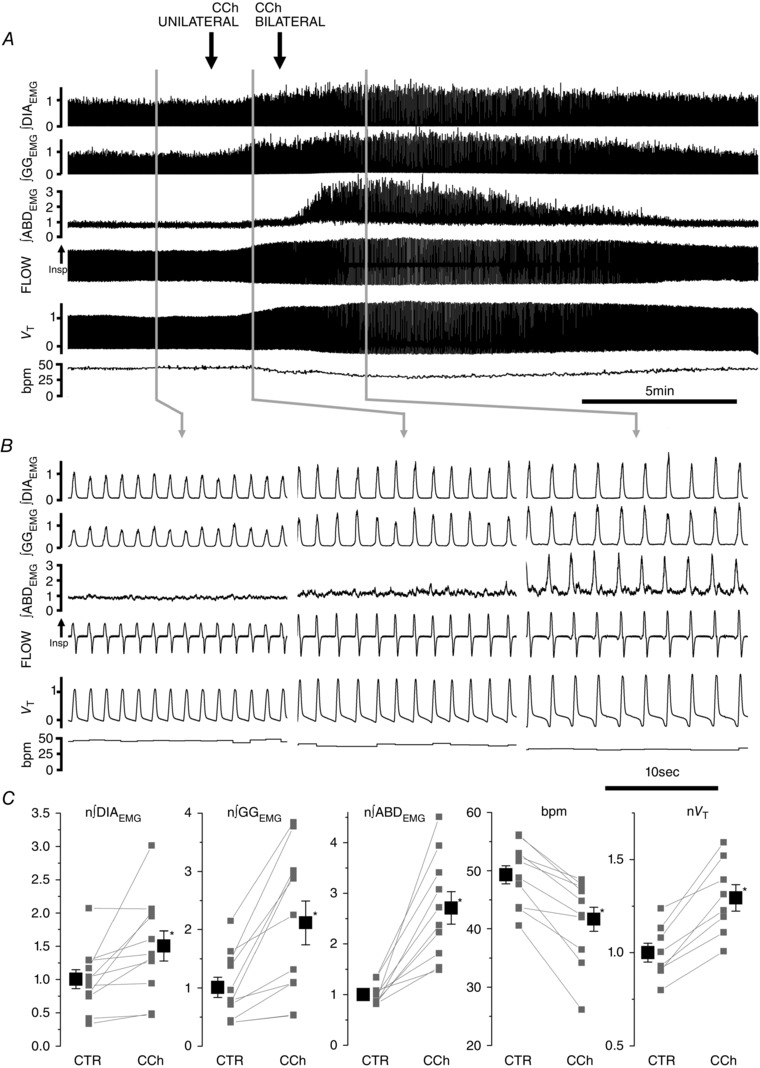
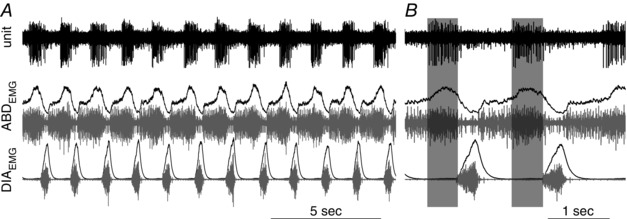
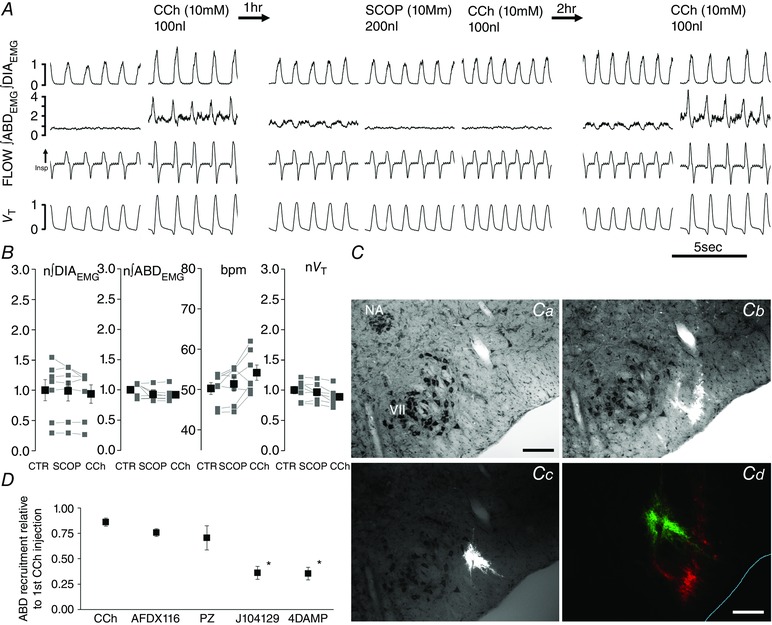
Abstract: Active inspiration and expiration are opposing respiratory phases generated by two separate oscillators in the brainstem: inspiration driven by a neuronal network located in the preBötzinger complex (preBötC) and expiration driven by a neuronal network located in the parafacial respiratory group (pFRG). While continuous activity of the preBötC is necessary for maintaining ventilation, the pFRG behaves as a conditional expiratory oscillator, being silent in resting conditions and becoming rhythmically active in the presence of increased respiratory drive (e.g. hypoxia, hypercapnia, exercise and through release of inhibition). Recent evidence from our laboratory suggests that expiratory activity in the principal expiratory pump muscles, the abdominals, is modulated in a state-dependent fashion, frequently occurring during periods of REM sleep. We hypothesized that acetylcholine, a neurotransmitter released in wakefulness and REM sleep by mesopontine structures, contributes to the activation of pFRG neurons and thus acts to promote the recruitment of expiratory abdominal muscle activity. We investigated the stimulatory effect of cholinergic neurotransmission on pFRG activity and recruitment of active expiration in vivo under anaesthesia. We demonstrate that local application of the acetylcholinesterase inhibitor physostigmine into the pFRG potentiated expiratory activity. Furthermore, local application of the cholinomimetic carbachol into the pFRG activated late expiratory neurons and induced long lasting rhythmic active expiration. This effect was completely abolished by pre-application of the muscarinic antagonist scopolamine, and more selective M3 antagonists 4DAMP and J104129. We conclude that cholinergic muscarinic transmission contributes to excitation of pFRG neurons and promotes both active recruitment of abdominal muscles and active expiratory flow.
Keywords: active expiration; carbachol; cholinergic modulation; parafacial respiratory group.
© 2016 The Authors. The Journal of Physiology © 2016 The Physiological Society.
Figures
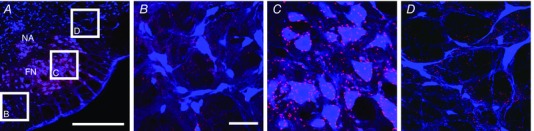
References
-
- Boers J, Kirkwood PA, de Weerd H & Holstege G (2006). Ultrastructural evidence for direct excitatory retroambiguus projections to cutaneous trunci and abdominal external oblique muscle motoneurons in the cat. Brain Res Bull 68, 249–256. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources