

On-chip, multisite extracellular and intracellular recordings from primary cultured skeletal myotubes
- PMID: 27812002
- PMCID: PMC5095645
- DOI: 10.1038/srep36498
On-chip, multisite extracellular and intracellular recordings from primary cultured skeletal myotubes
Abstract
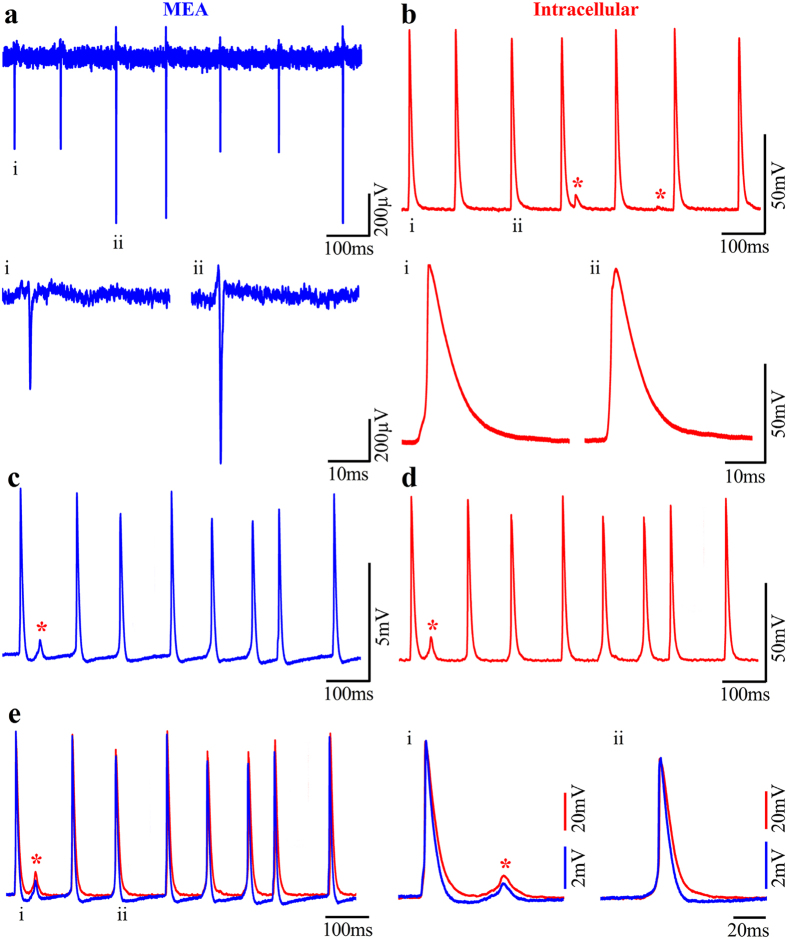
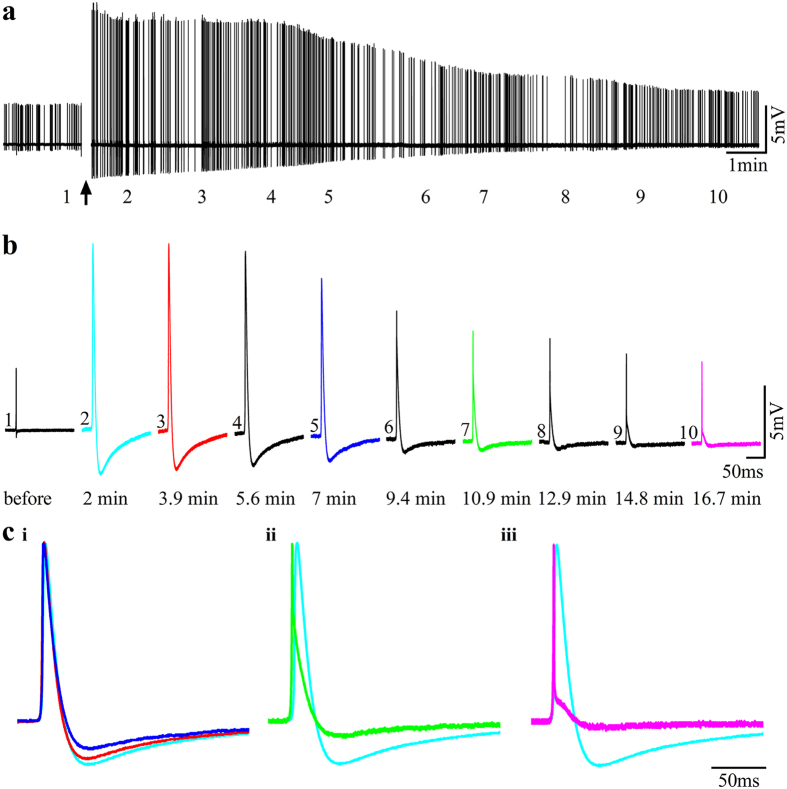
In contrast to the extensive use of microelectrode array (MEA) technology in electrophysiological studies of cultured neurons and cardiac muscles, the vast field of skeletal muscle research has yet to adopt the technology. Here we demonstrate an empowering MEA technology for high quality, multisite, long-term electrophysiological recordings from cultured skeletal myotubes. Individual rat skeletal myotubes cultured on micrometer sized gold mushroom-shaped microelectrode (gMμE) based MEA tightly engulf the gMμEs, forming a high seal resistance between the myotubes and the gMμEs. As a consequence, spontaneous action potentials generated by the contracting myotubes are recorded as extracellular field potentials with amplitudes of up to 10 mV for over 14 days. Application of a 10 ms, 0.5-0.9 V voltage pulse through the gMμEs electroporated the myotube membrane, and transiently converted the extracellular to intracellular recording mode for 10-30 min. In a fraction of the cultures stable attenuated intracellular recordings were spontaneously produced. In these cases or after electroporation, subthreshold spontaneous potentials were also recorded. The introduction of the gMμE-MEA as a simple-to-use, high-quality electrophysiological tool together with the progress made in the use of cultured human myotubes opens up new venues for basic and clinical skeletal muscle research, preclinical drug screening, and personalized medicine.
Figures






References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
