

Lipopolysaccharides-Induced Suppression of Innate-Like B Cell Apoptosis Is Enhanced by CpG Oligodeoxynucleotide and Requires Toll-Like Receptors 2 and 4
- PMID: 27812176
- PMCID: PMC5094738
- DOI: 10.1371/journal.pone.0165862
Lipopolysaccharides-Induced Suppression of Innate-Like B Cell Apoptosis Is Enhanced by CpG Oligodeoxynucleotide and Requires Toll-Like Receptors 2 and 4
Abstract
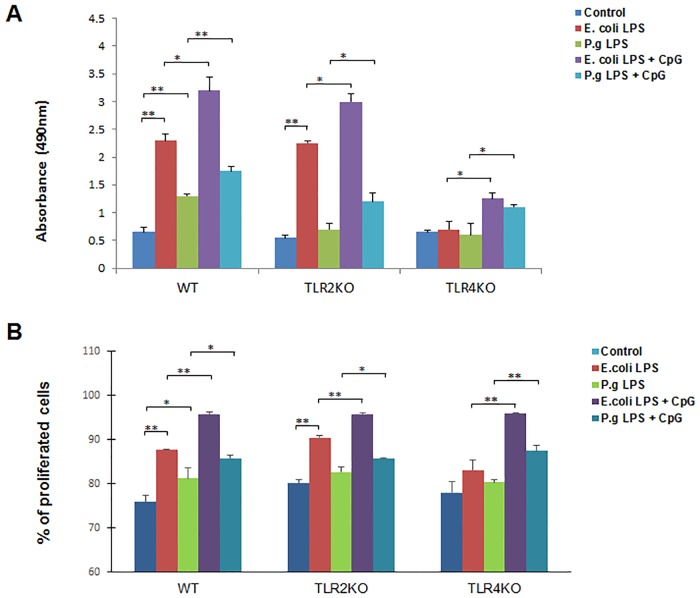
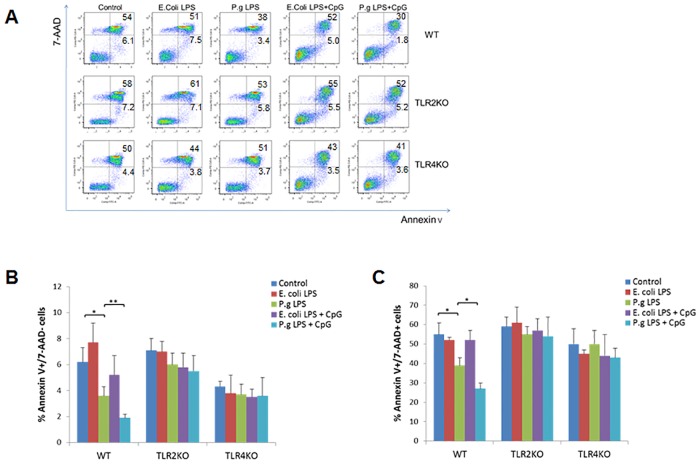
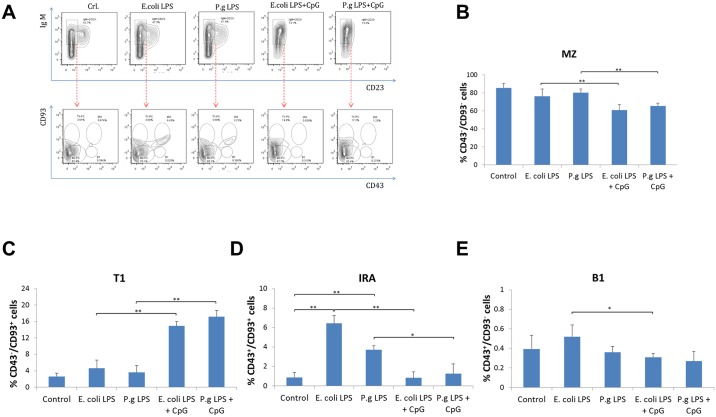
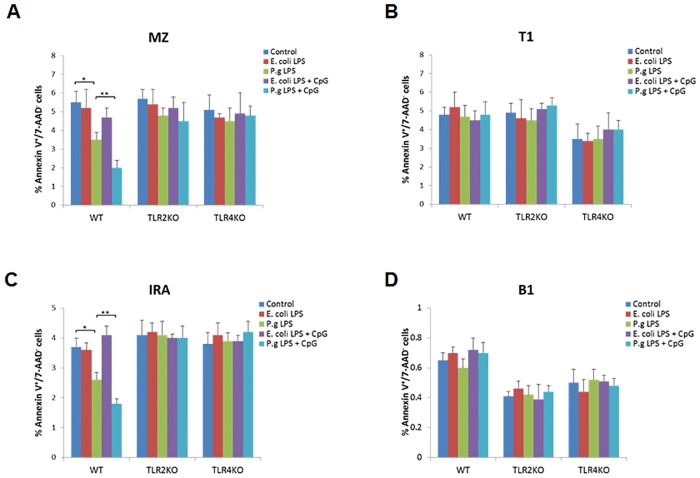
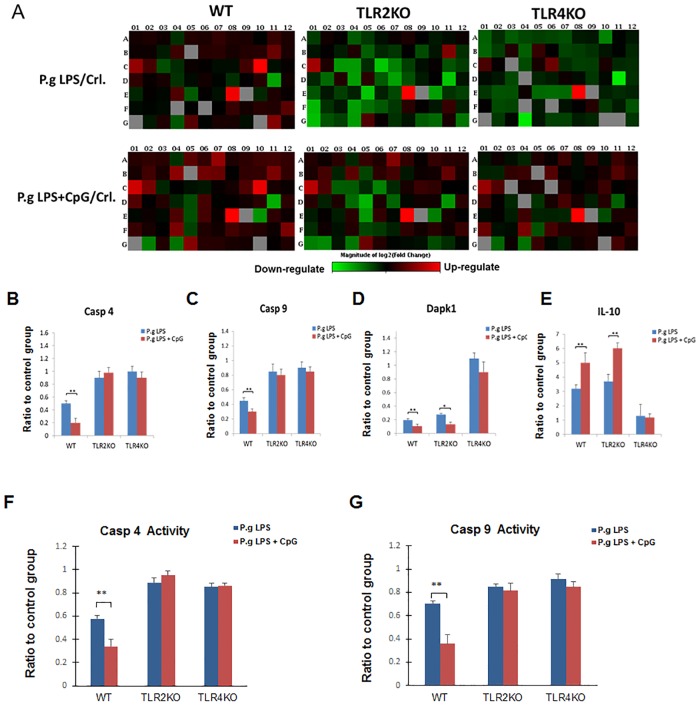
Innate-like B lymphocytes play an important role in innate immunity in periodontal disease through Toll-like receptor (TLR) signaling. However, it is unknown how innate-like B cell apoptosis is affected by the periodontal infection-associated innate signals. This study is to determine the effects of two major TLR ligands, lipopolysaccharide (LPS) and CpG-oligodeoxynucleotides (CpG-ODN), on innate-like B cell apoptosis. Spleen B cells were isolated from wild type (WT), TLR2 knockout (KO) and TLR4 KO mice and cultured with E. coli LPS alone, P. gingivalis LPS alone, or combined with CpG-ODN for 2 days. B cell apoptosis and expressions of specific apoptosis-related genes were analyzed by flow cytometry and real-time PCR respectively. P. gingivalis LPS, but not E. coli LPS, reduced the percentage of AnnexinV+/7-AAD- cells within IgMhighCD23lowCD43-CD93- marginal zone (MZ) B cell sub-population and IgMhighCD23lowCD43+CD93+ innate response activator (IRA) B cell sub-population in WT but not TLR2KO or TLR4KO mice. CpG-ODN combined with P. gingivalis LPS further reduced the percentage of AnnexinV+/7-AAD- cells within MZ B cells and IRA B cells in WT but not TLR2 KO or TLR4 KO mice. Pro-apoptotic CASP4, CASP9 and Dapk1 were significantly down-regulated in P. gingivalis LPS- and CpG-ODN-treated B cells from WT but not TLR2 KO or TLR4 KO mice. Anti-apoptotic IL-10 was significantly up-regulated in P. gingivalis LPS- and CpG-ODN-treated B cells from WT and TLR2 KO but not TLR4 KO mice. These results suggested that both TLR2 and TLR4 signaling are required for P. gingivalis LPS-induced, CpG-ODN-enhanced suppression of innate-like B cell apoptosis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Genestier L, Taillardet M, Mondiere P, Gheit H, Bella C, Defrance T. TLR agonists selectively promote terminal plasma cell differentiation of B cell subsets specialized in thymus-independent responses. J Immunol. 2007;178(12):7779–86. Epub 2007/06/06. . - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
