

BMP4 acts as a dorsal telencephalic morphogen in a mouse embryonic stem cell culture system
- PMID: 27815243
- PMCID: PMC5200901
- DOI: 10.1242/bio.012021
BMP4 acts as a dorsal telencephalic morphogen in a mouse embryonic stem cell culture system
Abstract
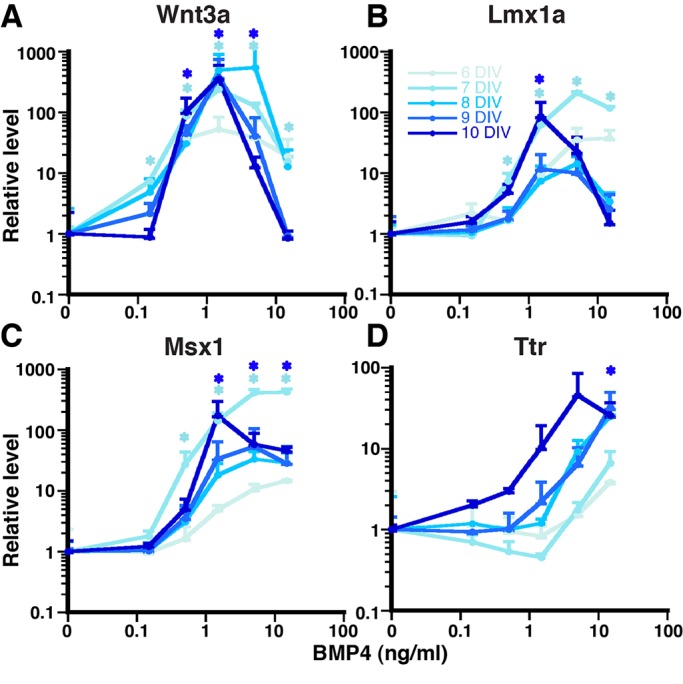
The concept of a morphogen - a molecule that specifies two or more cell fates in a concentration-dependent manner - is paradigmatic in developmental biology. Much remains unknown, however, about the existence of morphogens in the developing vertebrate central nervous system (CNS), including the mouse dorsal telencephalic midline (DTM). Bone morphogenetic proteins (BMPs) are candidate DTM morphogens, and our previous work demonstrated BMP4 sufficiency to induce one DTM cell fate - that of choroid plexus epithelial cells (CPECs) - in a mouse embryonic stem cell (mESC) culture system. Here we used BMP4 in a modified mESC culture system to derive a second DTM fate, the cortical hem (CH). CH and CPEC markers were induced by BMP4 in a concentration-dependent manner consistent with in vivo development. BMP4 concentrations that led to CH fate also promoted markers for Cajal-Retzius neurons, which are known CH derivatives. Interestingly, single BMP4 administrations also sufficed for appropriate temporal regulation of CH, CPEC, and cortical genes, with initially broad and overlapping dose-response profiles that sharpened over time. BMP4 concentrations that yielded CH- or CPEC-enriched populations also had different steady-state levels of phospho-SMAD1/5/8, suggesting that differences in BMP signaling intensity underlie DTM fate choice. Surprisingly, inactivation of the cortical selector gene Lhx2 did not affect DTM expression levels, dose-response profiles, or timing in response to BMP4, although neural progenitor genes were downregulated. These data indicate that BMP4 can act as a classic morphogen to orchestrate both spatial and temporal aspects of DTM fate acquisition, and can do so in the absence of Lhx2.
Keywords: Choroid plexus epithelial cell; Cortical hem; ES cell.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures




References
-
- Chizhikov V. V., Lindgren A. G., Mishima Y., Roberts R. W., Aldinger K. A., Miesegaes G. R., Currle D. S., Monuki E. S. and Millen K. J. (2010). Lmx1a regulates fates and location of cells originating from the cerebellar rhombic lip and telencephalic cortical hem. Proc. Natl. Acad. Sci. USA 107, 10725-10730. 10.1073/pnas.0910786107 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
