

Transcriptomic Characterization of SF3B1 Mutation Reveals Its Pleiotropic Effects in Chronic Lymphocytic Leukemia
- PMID: 27818134
- PMCID: PMC5127278
- DOI: 10.1016/j.ccell.2016.10.005
Transcriptomic Characterization of SF3B1 Mutation Reveals Its Pleiotropic Effects in Chronic Lymphocytic Leukemia
Abstract
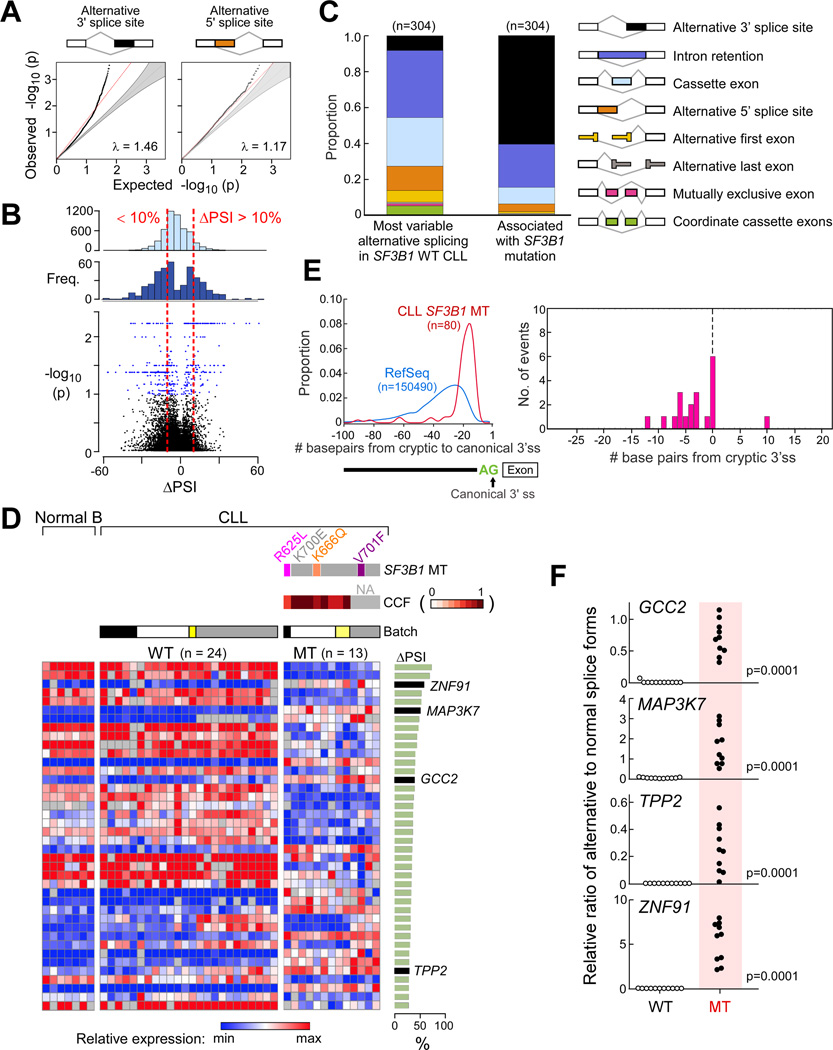
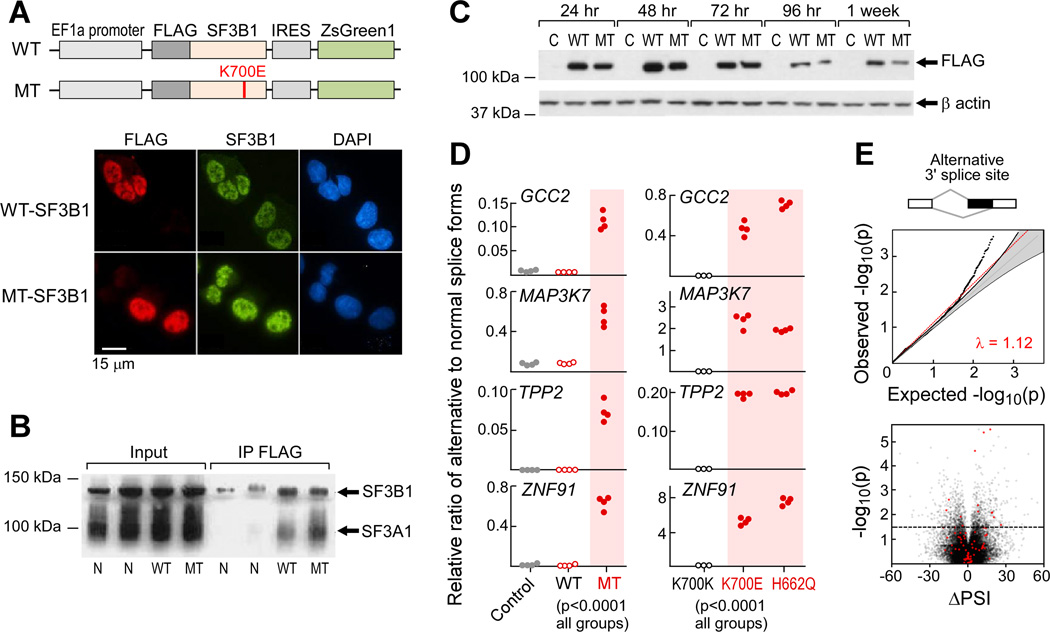
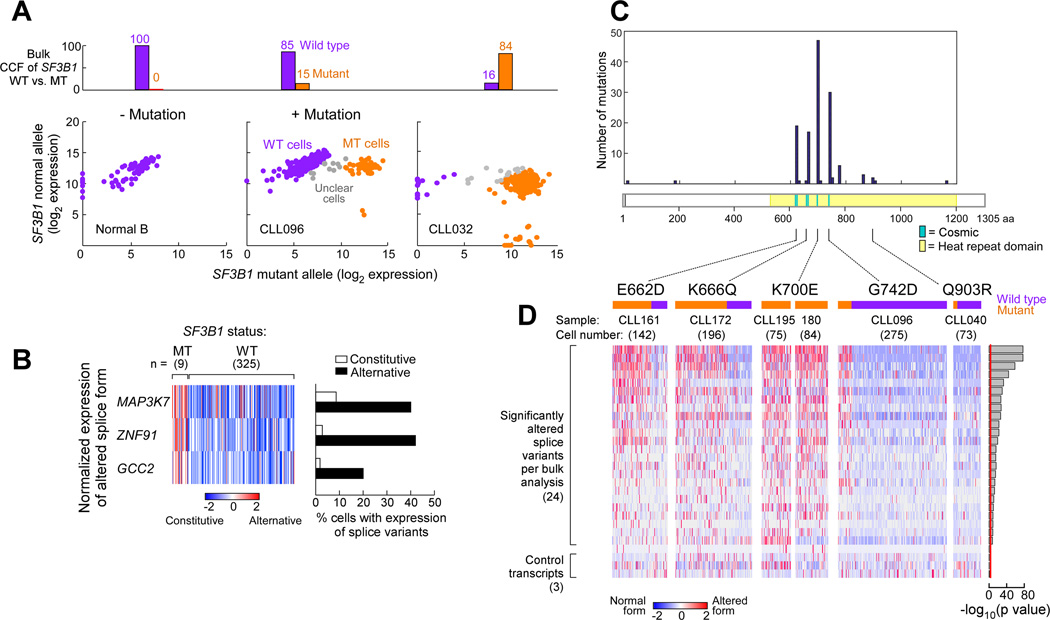
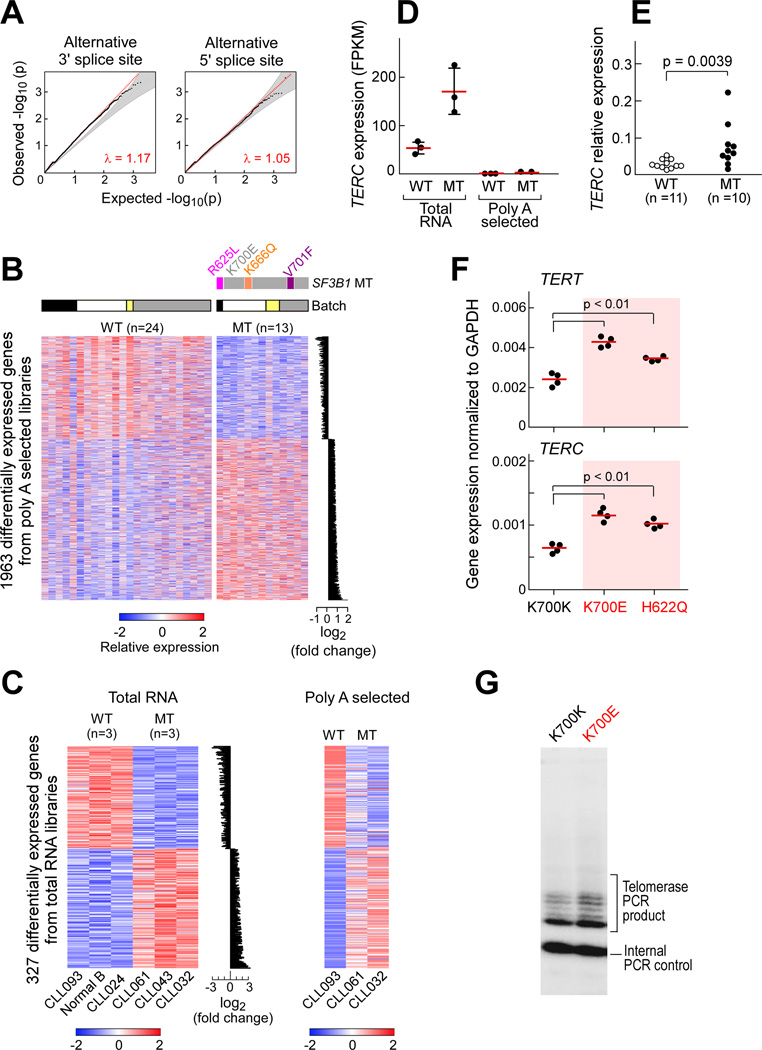
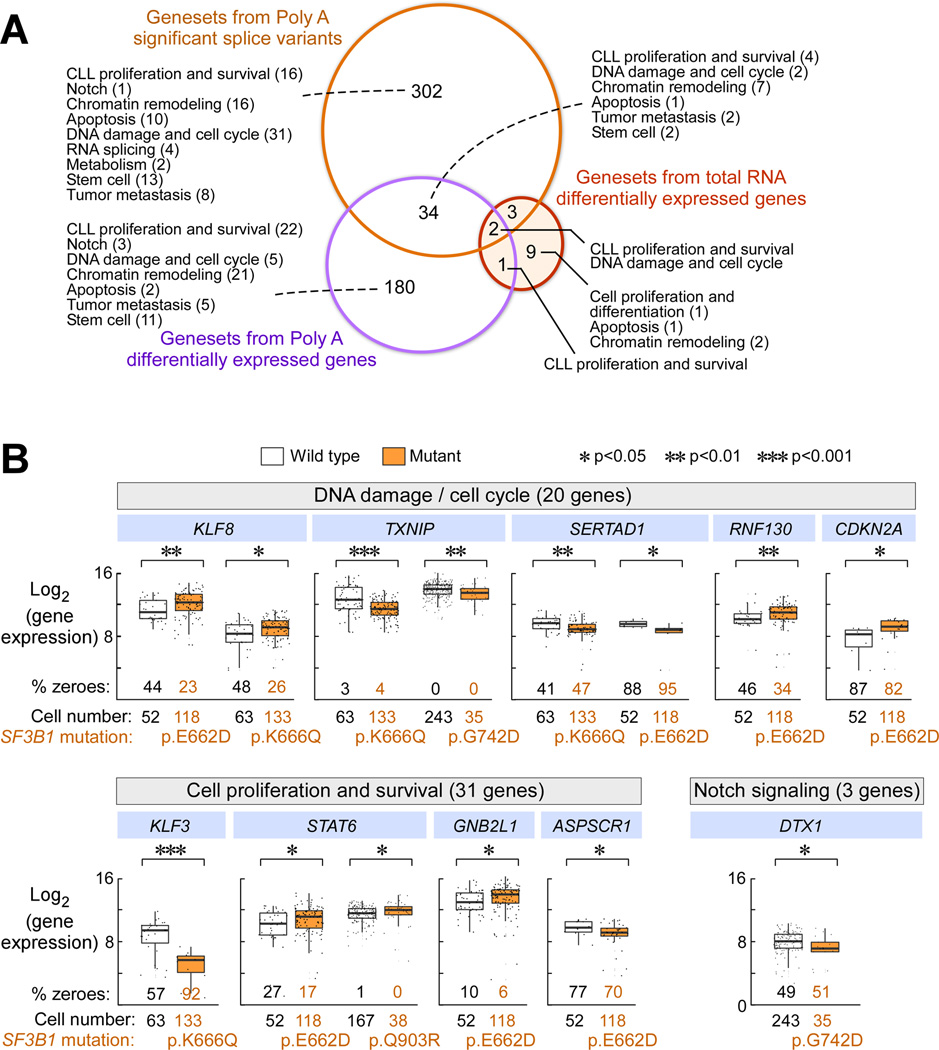
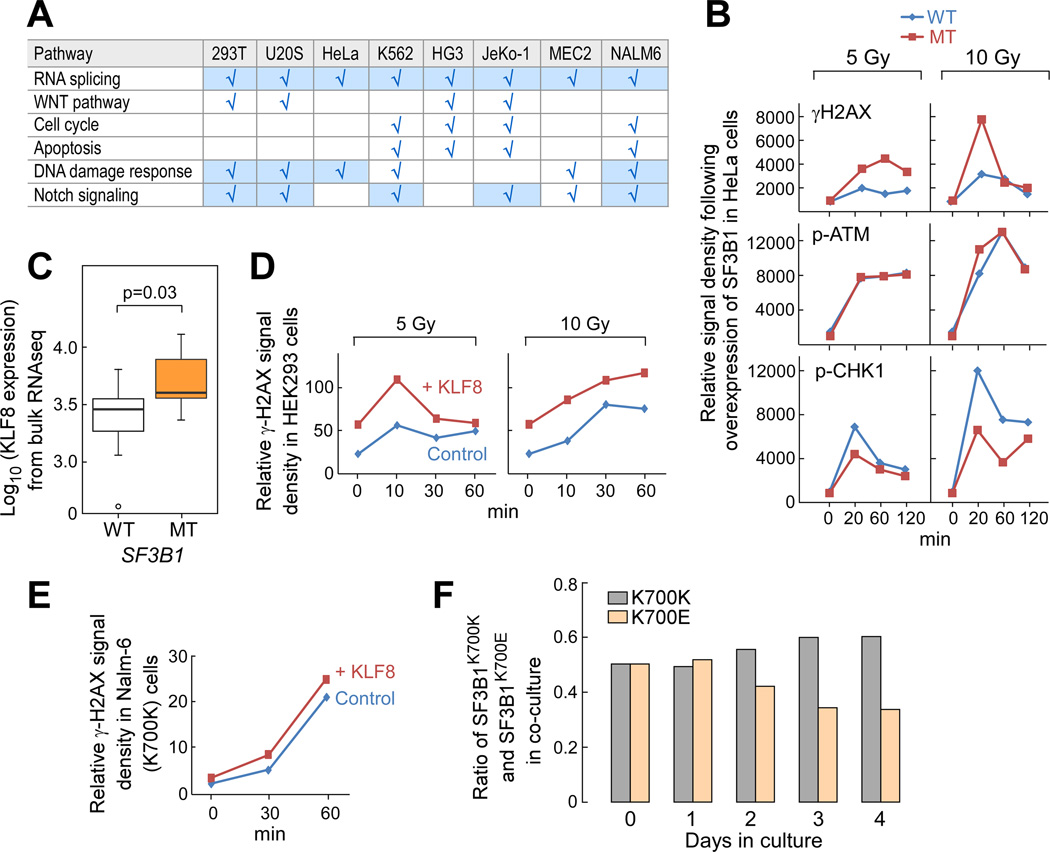
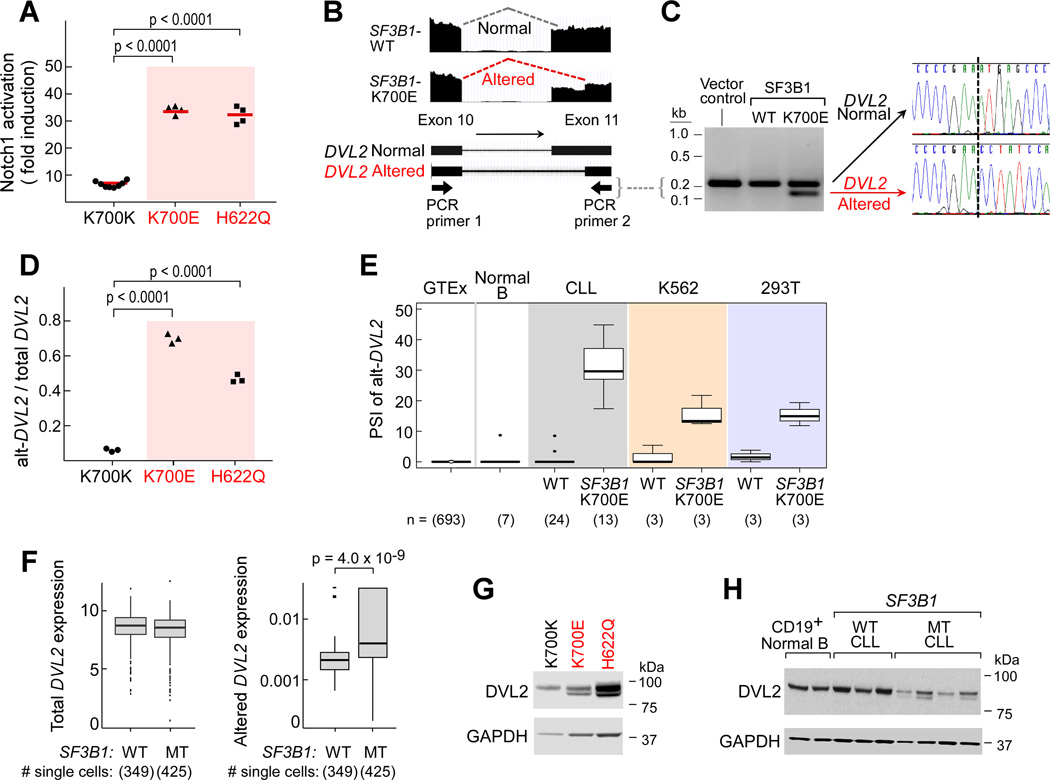
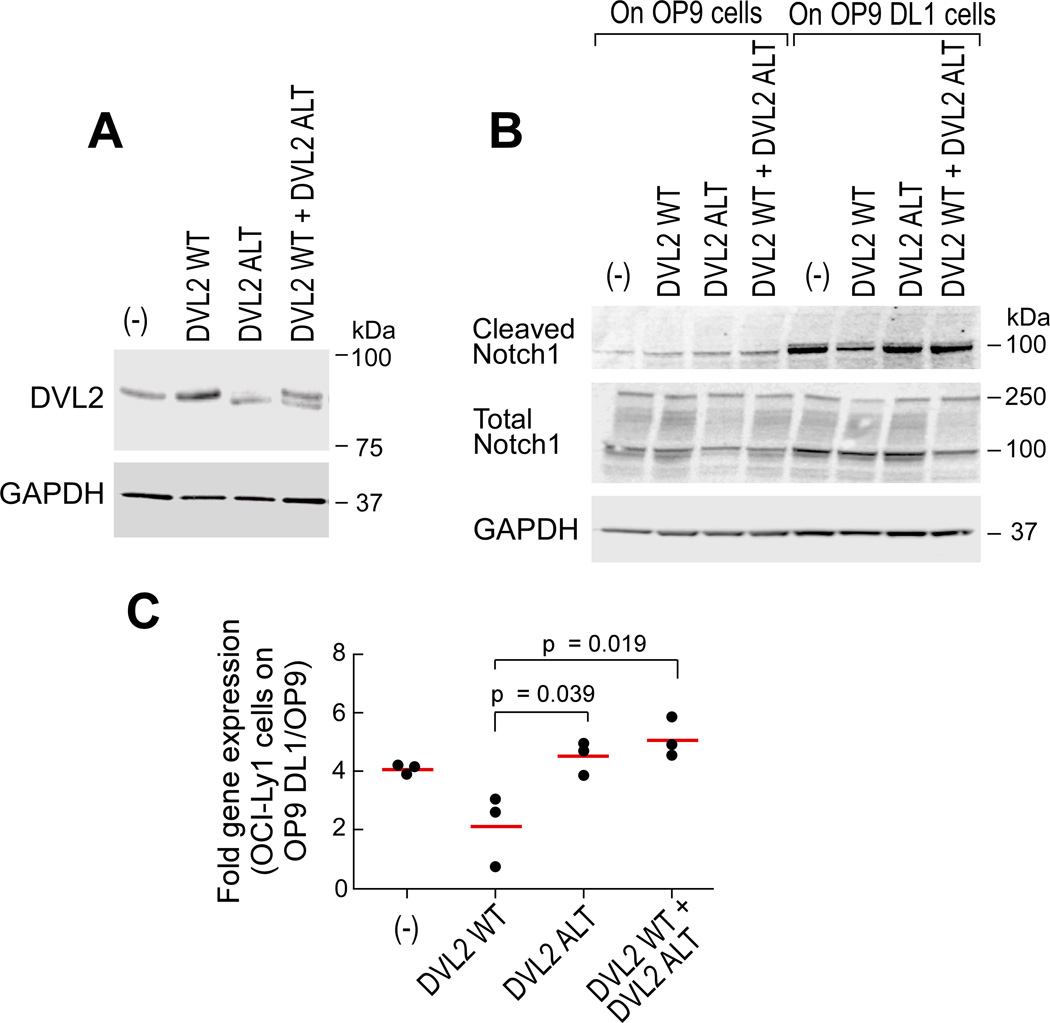
Mutations in SF3B1, which encodes a spliceosome component, are associated with poor outcome in chronic lymphocytic leukemia (CLL), but how these contribute to CLL progression remains poorly understood. We undertook a transcriptomic characterization of primary human CLL cells to identify transcripts and pathways affected by SF3B1 mutation. Splicing alterations, identified in the analysis of bulk cells, were confirmed in single SF3B1-mutated CLL cells and also found in cell lines ectopically expressing mutant SF3B1. SF3B1 mutation was found to dysregulate multiple cellular functions including DNA damage response, telomere maintenance, and Notch signaling (mediated through KLF8 upregulation, increased TERC and TERT expression, or altered splicing of DVL2 transcript, respectively). SF3B1 mutation leads to diverse changes in CLL-related pathways.
Keywords: CLL; Notch signaling; RNA sequencing; SF3B1; alternative splicing.
Copyright © 2016 Elsevier Inc. All rights reserved.
Conflict of interest statement
Michael Seiler, Silvia Buonamici, Peter G. Smith are employees and shareholders of H3 Biomedicine. Catherine J. Wu is co-founder and scientific advisory board member of Neon Therapeutics, Inc. All other authors have no conflicts of interest.
Figures
References
-
- Brooks AN, Choi PS, de Waal L, Sharifnia T, Imielinski M, Saksena G, Pedamallu CS, Sivachenko A, Rosenberg M, Chmielecki J, et al. A pan-cancer analysis of transcriptome changes associated with somatic mutations in U2AF1 reveals commonly altered splicing events. PLoS One. 2014;9:e87361. - PMC - PubMed
MeSH terms
Substances
Grants and funding
- U10 CA180861/CA/NCI NIH HHS/United States
- R01 CA182461/CA/NCI NIH HHS/United States
- F31 CA206236/CA/NCI NIH HHS/United States
- R01 HL131768/HL/NHLBI NIH HHS/United States
- R01 HL103532/HL/NHLBI NIH HHS/United States
- T32 GM007753/GM/NIGMS NIH HHS/United States
- R50 CA211482/CA/NCI NIH HHS/United States
- P01 CA206978/CA/NCI NIH HHS/United States
- P01 CA081534/CA/NCI NIH HHS/United States
- R01 CA184922/CA/NCI NIH HHS/United States
- R01 CA155010/CA/NCI NIH HHS/United States
- R01 GM043375/GM/NIGMS NIH HHS/United States
- R01 HL116452/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
